December 11, 1984
We seem to have fulfilled the classic image of the brain, which would consist of integration-differentiation by the centers, sensory-motor association, rational cutting. … If I tried to summarize what lived relationship we have with the brain, on the level of this image, it’s very simple, it is the brain tree, … that is, it’s a way of life, you have a tree in your head. This tree is this axis marked by rational points, rational breaks. And many people have lived with the idea and still live with the idea that their brain is a tree, that is, a centralized system, which means: center of integrations and differentiations, and sensorimotor associations, one expressing itself within the other and forming a circulation, a whole tree of the brain, a whole tree of the brain along these two axes. And, without wanting to analyze in detail at all, that poses all kinds of problems. I quote, for example, “What is the relationship between the centers and the elements? Assuming that the elements are brain cells, or nerve cells, the centers: what are these? Are they particular cells?” You sense here that there is a whole historical problem of localization exists – which I’m not pretending to address – of cerebral localization.
Seminar Introduction
As he starts the fourth year of his reflections on relations between cinema and philosophy, Deleuze explains that the method of thought has two aspects, temporal and spatial, presupposing an implicit image of thought, one that is variable, with history. He proposes the chronotope, as space-time, as the implicit image of thought, one riddled with philosophical cries, and that the problematic of this fourth seminar on cinema will be precisely the theme of “what is philosophy?’, undertaken from the perspective of this encounter between the image of thought and the cinematographic image.
For archival purposes, the English translations are based on the original transcripts from Paris 8, all of which have been revised with reference to the BNF recordings available thanks to Hidenobu Suzuki, and with the generous assistance of Marc Haas.
English Translation

Discussing three viewpoints on the brain (brain biology; lived relations with the brain; the brain as cinema) from which brain transformations have occurred, he pursues the biological, scientific review, and then from the lived, experiential perspective, he considers the classical, arborescent model of the brain with numerous theoretical references (cf. Jakobson on types of aphasia) allowing Deleuze to raise complications posed by this model. Then, shifting to the lived, experiential point of view, Deleuze links the arborescent model to another model from a classic perspective, that of interior and exterior milieus with details drawn from Simondon’s L’individu et sa genèse physico-biologique. With Simondon, Deleuze considers the brain’s topological structure, particularly questions of outside-inside, and then links thought of the Outside to the complementary sensorimotor axis. After the break, Deleuze outlines four lines of research for understanding cerebral linkages: the mathematical concept of Markov chains and the study of semi-accidental phenomena and mixtures of dependence and uncertainty; a second research line, within cosmobiology, of understanding the amplified fluctuation of enzymes and proteins from the perspective of the genesis of life; a third line of research (cf. Prigogine and Stengers), the thresholds at which the amplified fluctuations can be maintained in an orderly fashion; finally, the constitution of an internal milieu as an amplified fluctuation allowing the living creature, and even the brain (through hypotheses) to enter into aleatory relations with an external milieu (cf. cybernetician Pierre Vendryes). Deleuze answers the final question — how transmission occurs within the brain — with relinkages in independent series, suggesting finally the possibility of understanding cinema no longer as linkages but relinkages of independent series.
Gilles Deleuze
Seminar on Cinema and Thought, 1984-1985
Lecture 06, 11 December 1984 (Cinema Course 72)
Transcription: La voix de Deleuze, Laura Cecilia Nicolas (Part 1), Guadalupe Deza; correction Anselme Chapoy-Favier (Part 2), Sabine Mazé (Part 3); additional revisions to the transcription and time stamp, Charles J. Stivale
English Translation: Graeme Thomson & Silvia Maglioni
Part 1
Sorry to disturb you, but wouldn’t you say politeness is the most beautiful form of moral consciousness? Otherwise, it’s not that I think…
Anyway, so we’ve lost some time[1], but we’re going to make up for it because today we’re going to do a rather fuzzy session, since I’m not… I haven’t yet fully recovered. And this rather fuzzy session – and I have an idea that I’ll explain to you at the end of the session – concerns our last point, the last point of this program. I’d like us to get through it today, so I’ll go faster… no, not faster, not faster, no… I’ll just be a bit vaguer to save time. This last remaining point is the fourth aspect of mutation, the one that concerns the brain. So, we’ll have a chance to recap everything we’ve done this trimester, but I’m not going to recap just yet. I suppose you will recall, this was our fourth point, which would be the last one for us.
So, what has happened? “Mutation” may not even be the best word for it. It has happened in such a scattered, imperceptible way… but, as you know, because I’ve already said this, what interests me most are three points. What has changed from a certain point of view, from a triple point of view? First of all, from the point of view of the brain’s biology. This would be for the scientists, it’s one for the scientists, though I’ve never seen a case where such problems could be considered exclusively scientific. On the other hand, it’s not only from the point of view of the brain’s biology, but from that of a lived relationship with the brain. And finally, it would be from the point of view of cinema as brain or brain as cinema. I’m not saying that all cinema is brain, and all brain is cinema. But, if you remember what we’ve seen, we’ve seen certain things… for example, a cinema that could be very rudimentarily called a “cinema of bodies”. And, as I was saying, just as there’s a cinema of the body, there’s also a cinema of the brain.
So, my three points have to do with these three dimensions: potential changes, potential major shifts in the biology of the brain, potential major changes in our lived relation with the brain, if any, and potential changes in the cinema of the brain. And regarding this, as in the other cases, I try to oppose, though always very roughly, a so-called classical image, defining “classical” as vaguely as possible, and a so-called modern image, defining “modern” as vaguely as possible. This implies that if there is a modern cinema of the brain, for example, it does not proceed in the same way as what might be called the pre-war cinema of the brain. Just as brain models have greatly evolved in the sciences, this much everyone knows. But what we can all sense, even without knowing it, is that our lived relationship with the brain has also changed, at least in some cases.
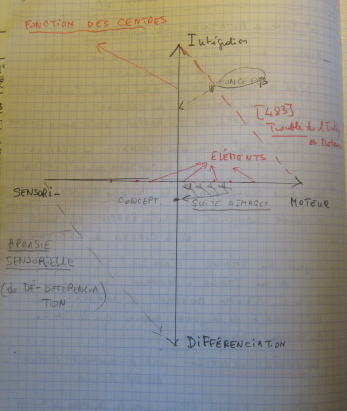
And so, I ask myself, what was the classical image of the brain? How could we present it? How could it be presented? Well, first of all from a scientific point of view. And, really, I don’t wish to say anything complicated in this regard. I would just say that in the scientific image of the brain, in the classical scientific image, it seems to me that there are two axes. One corresponds to the action of the centers, the other corresponds to the action of the elements. And it will already be quite a problem in neurobiology to know what the centers consist of and what the elements consist of, to know what their differences are, what the relations between them entail.
Suffice it to say that the centers that define cerebral areas, that define regions, cerebral areas, present themselves according to a double aspect: on the one hand… an indissociable aspect. They are “integrators”, the famous cerebral integrations. But they are also “differentiators”, differentiating areas. For example, we will differentiate between a projective area, which is to say one involving the projection of sense organs, a motor area and an associative area. But the action of integration is strictly indissociable from an action of differentiation. You see in the diagram on the board… where I’ve drawn my vertical arrow, my vertical arrow: integration-differentiation as functions of centers.
On the other hand, the horizontal arrow concerns the elements. Every element, every nerve element, every cerebral element is both sensory and motor. These are sensory-motor elements that enter into linkages or associations: sensory-motor associations or linkages, therefore sensory-motor associations or linkages that concern the elements, while integration and differentiation concern the centers. You will no doubt tell me that none of this surprises you… [Recording interrupted, 8:27]
… because in terms of the brain… we find the simplest givens of a biology of the brain, what we saw in the case of the classical image of knowledge. It’s as if there were an isomorphism between the model of knowledge and the model of the brain.[2]
Because, if you remember, when we were dealing with the model of knowledge in terms of another mutation, what did the classical image of knowledge consist of? One axis, which I called the concept axis, according to which there were concept integrations and concept differentiations and divisions, that is specifications. So, specifications and integrations which expressed the movement of the concept in knowledge. And on the other hand, there was another axis, which took the form of image associations. How did these axes relate to one another and circulate between themselves? It was very simple, and this was the model of knowledge. A concept could not be differentiated, that is, it couldn’t be divided, couldn’t be specified, without at the same time being externalized in a series of extendable images. But conversely, a series of images could not be extended without becoming integrated into a concept. Thus, we have a whole circulation taking place, which is the circulation of knowledge.
So, when I say that there exists an isomorphism in this brain model… indeed, the centers don’t differentiate without being externalized in extendable sensory-motor chains, while the sensory-motor chains are not extended without becoming integrated into the centers. So, this is normal, and it would be worrying if there were no longer this isomorphism between the old model of knowledge and the model of the brain. For reasons I don’t want to go into, I’d say that… no, I’m adding something here, but we’re still missing a point. If you’ve been following me, if you have any recollection of what we did previously, you will have realized that we’re missing a point. So, we have a duty here, but can we fill this gap? Ideally, in this old model of the brain, the cuts in the sensory-motor linkages, by virtue of the other axis, the vertical axis, would still have to be rational points.
In this way we would complete the classical image of the brain that consists of integration-differentiation through the centers… sensory-motor association and rational cuts. Now, the rational cut, I’ve already justified the first two points, but regarding the rational cut, the rational points… I haven’t been able to justify this yet. But if I could, then we’d all be very happy. So, I wanted you to feel this… if you did it would be a great joy, well, a small satisfaction anyway. But at least we’ve got the first two points.
I would say – and it’s not because of the schemas – but if I were to try and sum up the lived relation we have with the brain, in terms of this image, I would say it’s quite simple: it’s the tree of the brain. You might say, but it doesn’t look like a tree. Well, actually, it does look like a tree but we’ll see in what way. In other words, it’s a way of life: we have a tree in our head. This tree is our axis marked by rational points, rational cuts. And many people have lived according to the idea, and still live according to the idea, that their brain is a tree, that is, a centralized system. And by centralized system we mean a center of integration and differentiation, along with sensory-motor associations, each expressing itself in the other, and producing a circulation, a whole arborescence of the brain, a whole arborescence of the brain along these two axes.
Now, without wishing to analyze this in any detail, it poses all kinds of problems. For example, what is the relation between the centers and the elements? Assuming that the elements are brain cells, or nerve cells, what would the centers be? Would they be particular cells? This will be… you can see how there’s a whole historical problem of localization – which I don’t pretend to tackle now – of cerebral localization. But the problem is very, very complicated for a very simple reason. A great neurologist, a neurologist and psychiatrist, who had been greatly influenced by Bergson and whose name was [Constantin] Von Monakow, wrote an admirable book which I don’t think is read any more, a very, very fine book… M-O-N-A-K-O-W. He said: you understand, the mistake is to look for a spatial localization of the centers. The sensory-motor cells, yes, they have spatial localizations. But the integrating and differentiating centers have no spatial localization. And he proposed the idea of chronogenic localization, a chronogenic localization. So, he came up with some very complicated schemas. You can see why he was Bergsonian: he played with a kind of Bergsonian idea of durée inscribed in the living being and in the brain, and chronogenic localization was one of Monakow’s most complex ideas.
To give you an idea of the difficulties involved, I can say that a famous notion was put forward by a great neurologist named [John Hughlings] Jackson, the idea that brain diseases were in fact processes of disintegration, of dissolution. That is, the centers were damaged and, as a result, sensory-motor associations were unshackled from the centers’ integrating role. This is what he termed “dissolution”. And dissolution was the opposite of evolution. Evolution went from the elements to the centers.
What created the complications and made it so… Jackson said something very odd. He said that the centers are the least organized. In other words, the elements are much more organized than the centers, which seems strange. The centers, both integrators and differentiators, are much less organized. He explained very well what he meant by “less organized”. He meant that their organization continues throughout life. It was a way of linking up with Monakow’s chronogenic localization – not linking up, because it’s actually Jackson who precedes Monakow in history… – but in this way he anticipated the idea of chronogenic localization. The centers never ceased to establish and organize themselves throughout the whole of their life. So, they were less organized than the sensory-motor axis pertaining to the elements.
And again, you see how Jackson, in his theory of evolution and dissolution, was able to say that evolution proceeds from the most organized to the least organized, and dissolution the reverse, since the most organized is automatic: the automatic sensory-motor linkages. And so dissolution goes from the least organized to the most organized, just as evolution went from the most organized to the least organized. Nonetheless, it seems to me that in his conception he placed particular emphasis on the integrating role of the centers.
But here’s where I say it gets very complicated: in my view – and it’s just my view– there can be no phenomena of disintegration without correlative phenomena of “de-differentiation”. Therefore, patterns of dissolution become enormously complicated. There is no disintegrating action that is not accompanied by a de-differentiation, meaning a suppression of differentiation. And then, much more, perhaps, in phenomena of dissolution, there are disorders where disintegration is prevalent, and others where de-differentiation prevails. All of which considerably complicates our schema. So much so that the sensory-motor cells escaping from the centers disintegrate at the same time as they are de-differentiated, that is they lose their differentiation at the same time as they lose their integration. Hence, if I may jump now to a very famous problem, which has been quite decisive in brain mapping, this would be aphasia and the different types of aphasia. Aphasia, as you know, is a language disorder resulting from brain damage.
Now, traditionally, the great theory of aphasia distinguished – it went very far but it’s understandable, considering how complicated the axis already was – the great theory of aphasia distinguished between six types of aphasia from which, in the end, we could identify two major poles. And you’ll find a fairly detailed linguistic study of the two main poles in the work of [Roman] Jakobson. Which is a way of suggesting that Jakobson, in a certain way – implicitly, since he doesn’t claim to be a biologist – retains… [Recording interrupted; 22:57]
… but they can be reduced to two poles, two poles he studied in particular in his Essais linguistiques.[3] And these two poles will be self-explanatory, and we’ll see how they take into account our axis, or rather our two axes: a vertical axis and a horizontal axis. The two poles, he says, the two main poles of aphasia are first of all disorders that could be called “similarity disorders”, and then “contiguity disorders”. Similarity, he says, is the basis of what he calls “metaphor”, while contiguity is the basis of what he calls “metonymy”.
And if I briefly summarize these two disorders, the first, called similarity disorder… how does it manifest itself? The subject has retained the power to combine the elements of language. But what have they lost? They’ve lost… they have kept the power of combination, but they have lost the power of selection. They can construct a sentence but can no longer select between one element and another. What does, this imply? In a conversation, they respond very well, they can continue a conversation, or if presented with a fragment of a word or a fragment of a sentence, they can complete it very well. So, they’ve retained the art of combination. But they have a problem, which is obviously how to begin a sentence, it’s the initial word. Why is this? You can sense it. It’s because the initial word is something you have to select, you have to choose it. So now they are troubled. They are blocked. But once the phrase has begun, everything is again fine. There’s another moment that troubles them, however. It’s when they must select between two possibilities. For example, if they use the word “single”. For a normal person, the word “single” can be replaced with the word “unmarried”. But they can’t select, they can’t make this substitution. If you ask them what a single person is, they panic. They know how to construct a sentence with “single”, they know how to combine, but they no longer know how to select. And yet, as Jakobson would say, these are the two great poles of language: combination and selection.
What’s more, they can’t make the minimum selection of the type: A = A, or if you prefer, the minimum substitution, the identical substitution. Because if you say to them: “Say no”, they will reply: “No, I can’t”. It’s a seemingly paradoxical situation, since they can’t say “no”, but they say they can’t do it by saying “no”. You see, this actually concerns two very different axes of language. If we say to them: “Say no”, that is: “Repeat” – a substitution of the type A = A – they reply: “No, I can’t”, “No, I can’t say no”. The “no” they use in “No, I can’t”, is caught up in a combination, therefore it’s something they can do, whereas the selection of “no” and the minimal substitution of “no” by “no” is something they are unable to perform. It’s a type of aphasia, an aphasia of similarity or similarity disorder.
In contiguity disorder, the opposite is true. Similarity seems to be saved but composition is… selection seems to be saved but composition not in the slightest. In this type of aphasia, you encounter what linguists call “agrammatism”, that is, the dissolution of grammar. The sentence of this type of aphasic – you can see that it’s very different from the other case – the sentence of this type of aphasic ends up being a kind of fragmentation of words. What disappears are the links between words, and in particular inflexions. When they use a verb, this type of aphasic generally puts it, or would very often place it in the infinitive. Right. This time, as Jakobson says, similarity is saved. They can say the word “pencil” very well, they can make the necessary substitutions, but they can no longer make the combinations.
Well, to sum up very quickly – because I don’t want to dwell on this, I need to make up for the time I’ve lost – I’d say this: what is contiguity disorder according to Jakobson? Here, I’m speaking for myself, I’m no longer speaking according to Jakobson. Jakobson distinguishes very… I’ll come back to Jakobson quickly, he distinguishes perfectly between the two disorders by saying that the “contiguity disorder”, that is, when what is disturbed is the power of construction, is a disorder that relates above all to the message. Similarity disorder, on the other hand, is a disorder that primarily concerns the code. That’s easy to understand. When he’s more precise, because the interrelation between the code and the message is such that he indeed feels the need to be more precise, he tells us that similarity disorder is first and foremost a decoding disorder, and that contiguity disorder is first and foremost an encoding disorder. And in fact, anyone who’s studied even the least bit of linguistics will know all this.
As far as I’m concerned, I’d just like to draw the following conclusion: contiguity disorder is above all a motor disorder – you’re following my schema… “motor” is M, there, on the horizontal axis – it’s above all a motor disorder. In fact, it concerns the construction of spoken sentences and the combination of elements. And above all, it’s also a disintegrating factor. Why is that? As we’ve seen, it’s a disorder that concerns combination, and combination should be understood in two ways: combination of a linguistic unit by sub-units, by inferior units… by inferior units I mean, for example, phonemes, phonemes as sub-units in relation to morphemes – so a disorder of the combination of a unit by sub-units – and the combination by which a unit enters into superior units. It’s therefore a motor and integration disorder. Which is perfect for me, since it would allow me to draw a dotted line from I [Integration] to M which would represent this disintegrating motor aphasia. And motor aphasia was a common term in 19th-century aphasia theory.
The other disorder, similarity disorder, is essentially a sensory and differentiation disorder. Why? And what does it concern, exactly? As I said earlier, just as there are two levels of integration – the integration that a unit effects in relation to sub-units, and the integration into which it enters in relation to a higher unit – so too are there two aspects of differentiation. This can clearly be seen in language. There is a first aspect of differentiation, which we might call the discernment of distinctive features – I’m going quickly here because it doesn’t really matter, I’m only speaking for those who have studied a bit of linguistics – a phoneme is a carrier of a plurality of linguistic features, of distinctive features.
For example, when I say the word “cochon” [pig], I can extract the phoneme “ch”. Well, “ch” is, as linguists say, a cluster of distinctive features. Why is that? You’ve got “ch” rather than “t”, “cochon” rather than “coton” [cotton], you’ve got “ch” rather than “s”, “cochon” as opposed to “cosson”, you’ve got… Let’s look for other examples, what could it be? “Colon” [settler]… ah, yes, “ch” rather than “l”, “cochon” instead of “colon”. Each time you have… there isn’t a phoneme that doesn’t constitute a set of distinctive features. So, the first aspect of differentiation, I would say, is the selection of distinctive features, or rather… no, it’s the discernment, it’s the discernment of distinctive features. And the second aspect of differentiation is the substitution of two formulas that differ in one aspect and resemble each other… the substitution of one for the other two formulas that differ in one aspect and are similar in another.
It must therefore seem obvious to you that the kind of aphasia which presents phenomena of de-differentiation is also a sensory aphasia, while the aphasia which presents features of disintegration is necessarily a motor aphasia. So, my two types of aphasia, you see… I could draw a line going from I to M, that would be a disintegrative motor aphasia – and what I’m telling you now is no longer Jakobson, it doesn’t matter… but let’s not attribute it to him, he wouldn’t be happy – and the other aphasia S [Sensory] D [Differentiation], this would be the sensory aphasia operating by de-differentiation. That would be very complicated. What I’d like to show is that, in terms of the classical theory of aphasia, there are other types of aphasia that would allow us to fill in other blanks, but it wouldn’t give us much satisfaction. So, I’ll leave it at that for now. I hope you will vaguely remember this, or at least try to remember something of it, however vaguely.
I would say, well, basically, this would be the classical image of the brain. I used the example of aphasia because, on the one hand, I’m pleased to note that linguists… do you know what they’ve done? They’ve absolutely stuck to… in this new discipline, they’ve stuck to the oldest cerebral schema. So, I’m only mentioning the greatest among linguists. I’ve just tried to demonstrate this in the case of Jakobson, but for [Noam] Chomsky, I’d have to show it in detail, and it would take us three hours, but that’s all right. In Chomsky’s case, it truly is astonishing. Moreover, he admits it himself. When he made his return to Descartes, it wasn’t by chance. The cerebral model, that’s what interests me about linguists, knowing exactly what is the underlying cerebral model that they’re using. I believe that linguistics has an absolute need of a cerebral model and that it elicits it, it elicits and deploys it implicitly. So, when we turn to speak a little about these questions in terms of cinema with regard to linguistically-inspired semio-criticism, a principal question would be, for example, thinking of the very rigorous and important and highly interesting studies made by Christian Metz… What cerebral model does his conception of cinema imply?
And what strikes me about linguists in any case is the extent to which they cling to this conception of the brain: integration-differentiation of centers, sensory-motor association of elements. You might say, there’s no harm in it and I’m not saying that there is. There’s nothing wrong with it. It’s just that here we have a new discipline operating with an old cerebral schema. That’s all there is to it. I’m only rejoicing out of pure spite. Well, it strikes me that Jakobson’s conception of aphasia is very, very strong. You should read the book, because all these courses are designed to make you want to read this or that text. You’ll see, you’ll see for yourselves, but it’s no accident that he finds exactly the same schema of the various aphasias as that proposed by 19th century neurobiologists. And this concludes my first point.
I would say that what we have here is basically… basically, it’s the brain considered as a tree. Well, we live like that, or at least, we’ve lived for a long time like that, our fathers lived like that. It’s funny, imagine living… here, I’m talking about lived experience, I’m jumping to lived experience. I’m not saying much. But these are people who live with a tree in their head. There are a lot of people like that. And it’s no coincidence that the tree is both the model of the brain and the model of knowledge, from the tree of knowledge to the tree of the brain. Okay, short parenthesis.
What’s changed? I see two points of mutation, two points of mutation, but again, they overlap so much, it’s a very intricate, an extremely intricate business. I would also say that… to be precise here I have to go back… as well as being an arborescent system, I would say that this cerebral system is also a centralized system. It’s a centralized system because the whole vertical axis is based on the centers while the circulation is based on the interaction between these centers and the elements. Is that clear? So, it’s an arborescent, centralized system.
So now we’ll move on to the research into our mutation with the precautions I’ve taken. Bear in mind that it didn’t happen all at once, it occurred a little at a time. Let’s go back to the integration-differentiation axis, and here I would say that already in the oldest biological theories, and notably in the work of the great Claude Bernard, the problems of integration and differentiation have taken… have found themselves inexplicably tied up with another type of problem. And although the two were closely linked, they didn’t constitute the same problem. The other problem that intersected with the question of integration and differentiation was that of external and internal milieus. The more complicated and complex living beings became, the more they were composed of internal milieus. The stronger and more structured these internal milieus were, the more they depended on cerebral mechanisms and cerebral regulation. What’s more, organic differentiation and the organization of internal and external milieus intertwine at every moment. This means that the distinction between external and internal milieus promotes and accelerates the processes of organic differentiation that it, at the same time, implies. Here, too, there is complete interaction between differentiation and the constitution of internal and external milieus.
So how do integration and differentiation come about? Among other things, they arise through relative levels of exteriority and interiority. Why do I say relative? Because, at first sight, there are no absolute external and internal milieus. What from one point of view is an internal milieu becomes an external milieu from another. It all depends on which organic unit you take into consideration. You can, of course, take the organism as a whole, but even that will present difficulties. If you take the organism as a whole, you’ll tell me it’s not complicated: the internal milieu is what is inside. What is it that defines an evolved organism? Precisely the fact that it has constituted an internal milieu. What does it mean to constitute an internal milieu? It means constituting a milieu that no longer depends on variations in external milieus. A self-regulating milieu. For example, the heat of the blood becomes independent of variations in the external milieu. You have an internal milieu.
If you take the organism as a whole, you might say, Well, it’s not complicated; the internal milieu is everything that’s inside. But there’s no guarantee. We can’t be sure. What would you say about the stomach? Is it an internal milieu? Yes, it’s an internal milieu, but up to a certain point. In relation to what is it internal? Because in relation to something else it may be external. From another point of view, an internal milieu can be an external milieu, which we will call an external milieu annexed to the organism. All the more so for the intestine. The intestinal milieu is an internal milieu. But from another point of view, it plays the role of an annexed external milieu. So much so that intestinal waste is indeed discharged into the external milieu.
There’s a text by [Gilbert] Simondon in his very fine book, L’individu et sa genèse physico-biologique [The Individual and its Physical-Biological Genesis] [4] that emphasizes this point well. Here is what he says, and he says it better than I can, so I might as well read it to you: “There are several levels of interiority and exteriority; thus, a gland with internal secretion discharges the products of its activity into the blood or into some other organic liquid: in relation to this gland” – in relation to the gland with internal secretion – “the internal milieu of the general organism is in fact a milieu of exteriority. Similarly, the intestinal cavity is an external milieu for the assimilating cells that ensure selective absorption along the intestinal tract. […] The space of the digestive cavities constitutes an exteriority in relation to the blood that irrigates the intestinal walls. But the blood is in turn an external milieu in relation to the glands with internal secretion, which pour the products of their activity into the blood.”[5]
So that’s all I want to say. And here too, I don’t have time to develop because… The hypothesis I’d like to advance is that processes of organic integration and differentiation are strictly inseparable from the distinctions between internal and external milieus – these distinctions remaining relative, strictly relative, which means that a milieu said to be internal from one point of view will be external from another point of view, and that there will be a whole chain of internal and external mediations. These distinctions between internal and external milieus remain relative as long as we relate them to the integration-differentiation axis. So the integration-differentiation axis passes through these distinctions, which are relative to it. Okay, that must be very clear because…
All right, then. But that doesn’t prevent us… Simondon adds in some very, very strange passages that I nonetheless find quite admirable… if we refer to the state of the simplest organism, the simplest organism… this is all quite tedious, but thanks to Simondon the conclusion will be beautiful. If we look at the simplest organism, what happens? The simplest organism, undifferentiated and unintegrated, enjoys absolute interiority and exteriority, which are distinguished by what? By a polarized membrane, the simplest expression of life. There’s the outside and there’s the inside. In other words, the simplest organism is arranged according to an absolute outside and inside. As organisms become more complex, that is, as they integrate and differentiate, they pass through relative levels of interiority and exteriority.
That leaves the third conclusion, which you can work out for yourself: the brain restores, in its own way, an absolute outside and inside which is like the limit of all relativity of the external and internal milieus. What does this mean for the brain? Well, the brain will bring into co-presence an absolute inside and an absolute outside. What do I mean by co-presence? They will be in contact, in contact without any distance between them. In this way, outside and inside are no longer relative. The brain restores an absolute outside and inside, in other words, that which pertained to the simplest organism. It restores it in its own way. It brings into contact an absolute outside beyond all external milieus, which will undoubtedly be like the horizon of all external milieus, and an absolute inside, beyond all internal milieus, which will undoubtedly be – how shall I put it, poetically speaking – the most bottomless, or the deepest, of all internal milieus.
The brain will bring into contact an outside deeper than all external worlds, an inside – you can see where I’m going with this, if you remember what we did earlier – and an inside deeper than all internal worlds and milieus. How does it produce this contact, a contact without distance between outside and inside? The elementary organism required a distance between its absolute interiority and its absolute exteriority. What does contact without distance mean? That’s what Simondon explains in these wonderful passages. It means that the brain has a topological structure… [Recording interrupted; 58:10]
Part 2
… The brain has a topological structure that ensures the co-presence of an absolute outside and inside. You might ask what this is. What does it mean? Well, okay… There are just two things I’d like to point out here. As far as I know, even neurobiology has nothing to say on the matter, and it took a philosopher with great expertise in biology, technology and cybernetics to remind us of it, because I have the impression that cybernetics is very much concerned with this question. In the end, as Simondon says, the brain has no Euclidean interpretation. The brain cannot be interpreted in Euclidean space. Integration-differentiation can be interpreted in Euclidean space, and so too can relative internal-external milieus. But what does the brain imply? A topological space.[6]
But let me quote an interesting text here: “The development of the neocortex in the higher species takes place essentially through a folding of the cortex: it’s a topological solution, not a Euclidean solution. Ultimately…” – I think this text is very fine, it’s not important if you don’t understand much – “Ultimately, we shouldn’t talk about projection regarding the cortex” – he’s referring to what I told you about earlier: the areas of sensory projection, that is, the projection of the sense organs into cerebral areas – “Ultimately, we shouldn’t talk about projection regarding the cortex” – and, indeed, there isn’t a biologist in the world who doesn’t know that the supposed projection of sense organs onto a cerebral area are crude, senselessly crude schemas – “we shouldn’t talk about projection regarding the cortex, although there is, in the geometrical sense of the term, projection regarding small regions. We should rather speak of a conversion of Euclidean space into topological space. The basic functional structures of the brain are topological; the bodily schema converts these topological structures into Euclidean structures through a mediate system”.[7] By mediate system he means through the mediation of relative internal and external milieus.
And what does that mean? What does it imply? It means to bring into contact or into co-presence an interior that is deeper than all relative internal milieus, and an exterior that is deeper than all relative external milieus. That would be it. That would be the topological function of the brain. You see… this allows him to say: “The non-topological structure of integration and differentiation, the non-topological structure of integration and differentiation appears as a means of mediation and organization”– which is to say, of mediation and organization thanks to the relative internal and external milieus – “appears as a means of mediation and organization to support and extend the first structure” – that is, the topological structure – “which remains not only underlying, but fundamental.”[8]
Now this makes me very happy because it confirms, though using completely different means…We’ve been turning round and round this question – when I recap on all this next time, you’ll see, you’ll remember everything we’ve seen about this mutation of thought, insofar as it becomes a thought of the Outside, but of an Outside that is deeper than the external world, and which is also a thought of the inside, but an inside deeper than any internal milieu. And here, in terms of the brain, we find exactly the same problem, at the level of a topological relation, between an absolute outside and an absolute inside. In concrete terms… you will no doubt tell me that you’re begging for something more concrete, but what does that mean? It means something very simple. It means: the more an organism is evolved and subjected to a structured brain, the more the totality of its past, that is, the absolute inside – the absolute inside being precisely the All of the past – the more the totality of its past is brought into topological contact, without distance and without delay – topology ignores distances – without distance and without delay with the opening out of the future, that is, with the absolute outside.
Understand what this means. But that’s the way it is with living beings. It’s even what distinguishes the living from the crystal. When a crystal grows, what matters? It’s the molecular layers: they’re relatively internal and external. In other words, the molecular layer that has already formed is called the internal layer, while the molecular layer that is in the process of being formed is called the external layer. Well, you can empty the crystal of most of its internal substance, and it won’t change a thing. You cannot prevent it from growing. On the contrary, you cannot empty an organism of its internal substance. Its internal substance concentrates and condenses the whole of its past. And it’s this all of its past, the Whole of this past, which is in immediate contact, that is, topologically, which is – how would we say it in topology? – which is in the vicinity, independently of any distance, which is in the vicinity of the absolute Outside, that is, of the horizon of the outside world.
Therefore, we would have to say that the brain topologically reconstitutes the ordinary conditions of the simplest organism. But it is precisely in order to find them topologically that the highest complexity is required. Co-presence – even if we will have to justify it later, but we have all year to do that, don’t we? – co-presence that I could say, I could say from that point on the brain, being topologically interpreted – and it must be topologically interpreted, it can only be interpreted topologically – the brain would present itself as “contact without distance” or “co-presence” of outside and inside, of empty and full, of past and future, of front and back, and so on.
You have to hold onto these formulas, of the full and the empty. You can see that here I already have a cinematographic afterthought. What is fuller than an empty image in cinema? An empty space in [Michelangelo] Antonioni is a full image, an empty sky in [Jean-Luc] Godard is a full image. But full of what? [Recording interrupted; 1:07:19]
… no longer defined along an axis, it will no longer be defined… I don’t mean that it’s mutually exclusive, it’s not that one is false and the other… But the fundamental thing will no longer be an axis of integration-differentiation, it will be a co-presence of an absolute Outside and an absolute inside, of void and plenitude, of a front side and a reverse side and so on. If you experience your brain in this way – it’s not even necessary that you understand… – but if you experience your brain in this way, if you experience it topologically, and no longer in terms of integration and differentiation, well, in that case you will no doubt feel that you no longer have the same relationship with it, wouldn’t you say? Is it possible to experience it in such a way? Well, that’s something we’ll have to see… But it’s not easy. Do you insist on living according to a form of integration-differentiation? If so, are the gaps the same in both cases? What will the implications be for our conception of aphasia? But let’s not get ahead of ourselves.
I would say, and you must sense it yourselves, that this is not sufficient. A second, a second small mutation is required. A second small mutation because, otherwise… Fortunately, this time it will be much simpler, because the first one was quite complicated. For those who want to look further in this direction… the relevant pages are… from 257 onwards…[9] So, now I would say… you see how I’ve just dealt with my vertical axis… and now we’re going to replace my old vertical axis of integration-differentiation will a topological axis. Do you follow me? And, as I said, I need a second mutation because we now have to deal with the S – M axis, the sensory-motor axis, the associative axis. So, this is something that works by itself.
How does the sensory-motor axis present itself? It takes the form of a brain cell, or nerve cell, called a “neuron”. A neuron – I’ll give it to you, for those of you who haven’t… let me remind you of the basic vocabulary… I’m using it at the level of the most basic terms – a neuron is a cell, a brain or nerve cell. This cell has extraordinary branching extensions. These extraordinary branching extensions are called “dendrites”, D-E-N-D-R-I-T-E-S. You have your nerve cell, your neuron, and then it has extensions called dendrites. So, dendrites are branching extensions that receive nerve impulses. To give you an idea of just how complicated this is, it’s incredible how many dendrites a neuron has. It has a huge number of dendrites. But there’s much more to it. There are some that extend very far, that extend extremely far. This is where things get complicated.
And then, finally, you have the third major element, the “axon”, the axon, the axon: A-X-O-N. The axon is also an extension. But this time, it’s a single extension of the neuron, a single extension that branches off at the end – so it can branch off a lot – which branches off at the end and transmits the impulse. To what? The simple answer would be: to the dendrites of the following neuron, the next neuron. It all depends. Or it might not be to the next neuron, it’s not fixed since there are dendrites that extend considerably. So, you must realize, then, that this isn’t just automatic, that any one of your neurons may sometimes be subject to very distant impulses. There’s a multitude of impulses being triggered at the same time. It’s… it’s a whole system.
For a long time, the classical image of the brain was that all of this formed a continuous network. But it soon became evident that this network was not continuous, in the classical sense of the word, but was riven with cuts. These cuts were established, for example, between the end of the axon of neuron A and the end of the corresponding dendrite of neuron B. Remember? The affected dendrite receives the impulse, transmits it into the cell, into the neuron, which sends it to the axon, which passes it on to the dendrites of the next cell. However, between the end of the axon of cell A and the dendrite of cell B there is a cut. The network is not continuous.
But for a long time, the cut was conceived, so to speak, in an electrical manner, that is, from one edge of the cut to the other, the transmission of the impulse was electrical. This cut has a name, it’s called a “synapse”. A synapse is the communication, in the simplest case, between the end of an axon and the beginning of the dendrite of the next cell. So, we can literally say that a synapse is a cut-point. That’s why, very often in textbooks, you will see the synapse defined as a junction point. It’s the point where two neurons meet. It’s a point or a cut. We shouldn’t be surprised by this, given what we’ve seen and done previously. So, we’ll say it’s a cut-point.
So, as long as there was transmission, the hypothesis of electrical transmission, everything was fine. Everything was fine, or relatively fine. There was no longer a continuous network, but the impulse could nonetheless easily cross the cut-point. Why was this? Because, at the moment of passage of the impulse, the hypothesis was that the two cell membranes, meaning the dendrite membrane and the axon membrane – or rather the reverse, the axon membrane and the membrane of the following dendrite – came together, became adjacent, became adjacent… or even, according to a particularly audacious hypothesis, formed a single membrane. Whether they are adjacent or form one and the same membrane, here again we have, we have every reason to rejoice because in my view, by virtue of what we have seen previously and without in any way forcing the issue, we are entitled to say that in this case we have a rational cut-point.
So, without forcing the issue, without resorting to a facile mathematical metaphor – because, if you remember, this was our starting point, you have to remember this, otherwise it’s a disaster. What we called a “rational cut-point” on a straight line, on a continuous straight line, was a point that operated through division into two sets, but such that this point formed either the end of the first set or the beginning of the second set, meaning that it pertained to one or the other of the two sets, or else belonged to both at the same time… If someone tells me that in an electrical transmission, the two membranes become adjacent, I would say that one marks the end of the first set and the other marks the beginning of the second set. This is a typical cut-point. The synapse is a rational cut-point. And if someone says to me that a single membrane is formed, I say, Fine, the cut-point pertains to both sets. It’s a rational cut-point.
This gives me the third aspect of the classical image of the brain: a process of integration-differentiation exercised in relation to, second aspect, exercised in relation to sensory-motor linkages, the points – and this is the third aspect – the cut-points of these linkages being rational points, meaning that the synapses are rational points. I’m laughing to myself a bit because here I am subjecting you to a very unpleasant regime that goes from rudimentary mathematics to rudimentary biology, but I hope you will understand that all this is just a way of fashioning philosophical concepts, it’s just material… Do you follow me? I’d say, here are the three aspects, including the one I was missing. It’s curious how all this can be a source of deep satisfaction. We now have a third aspect, the one we were missing, and now we have it. We’ve reintroduced rational cut-points in the brain, or into the brain-tree. And so now this gives us an overall picture of the brain as tree.
So, this great mutation no doubt occurred when doubts began to arise, not about the existence of a continuous cerebral network – that question was settled, the existence of synapses was proven, nobody could argue about that – but about the way synapses actually function. And when the doubts multiplied about the electrical transmission hypothesis, a completely different hypothesis emerged: the idea of chemical transmission. What does this mean? It means that – you’ll immediately understand the difference – when the impulse arrives at the end of the neuron in cell A, neuron A… well, there’s no adjacency of membranes, no membrane unity. It finds itself before a cleft, it has a cleft to cross. Imagine – now you’re thinking things are starting to liven up – imagine that our brain will be populated with… well with what? It will be populated by millions and millions of little clefts, synaptic clefts. Well, you’ll tell me: It was already… there were already cuts. Well, okay. What’s going on here? In the words of a very fine author, there is a “critical gap”. Between two neurons, a critical gap.[10]
How does the impulse… if the impulse lacks the power to transmit itself electrically, how will it manage? In science, to transmit something electrically means by way of charged ions, right? If this is not the mechanism by which the synapse functions, then how does it work? Well, we might begin to suppose – and we have good reason for doing so – we might begin to suppose that the impulse, when it arrives at the end of the neuron, will release something that belongs neither to it, the axon of A, nor to the other, meaning the dendrite of B. It will release a chemical substance more or less similar to hormones: a chemical mediator, a chemical mediator. But where does this chemical mediator derive from?[11]
Fortunately, with advances in microbiology, bizarre vesicles, small vesicles, have been identified in the synaptic cleft. So, you see? From there, we can assume that these vesicles contain the chemical mediator and that when the impulse reaches the end of the axon of neuron A, when the impulse reaches the end of its path, the vesicle will release a quantity of the chemical mediator. This is getting more and more interesting, because if it is these vesicles that are releasing it, the release of the mediator will itself not be continuous. There will be discontinuities everywhere. A vesicle will contain a very small quantity of neurotransmitter, a chemical mediator of the hormonal type. It will release it. So, the chemical mediator will be released in discontinuous quantities corresponding to the different vesicles that release it. This is why neurobiologists speak of “quanta”. The chemical mediator operates in discontinuous quanta. And it is via the chemical mediator that the impulse will pass from neuron A to neuron B.
What does this tell me? I don’t need to… The cut-point, the synapse, remains a cut-point. This cut-point determines two sets: neuron A, with its axon and dendrites, and neuron B, with its dendrites and axon. The cut-point determines two sets. Only, it’s no longer part of either of the sets it determines: it’s autonomous. This is not a metaphor, it’s literally what we call an “irrational cut”, which is to say, a cut that determines two distinct sets, and yet belongs to neither of the two sets. I’d say that in the case of chemical synapses… whilst in the case of electrical synapses we were dealing with rational cut-points, in the case of chemical synapses we are faced with irrational cut-points.
And then… and then, it’s the whole cerebral regime that passes under a new regime that is literally – we’ll see where it leads us – a probabilistic regime. In other words, under a probabilistic regime, what are the probabilities that a message or an impulse arriving via the axon of A will cross the critical gap, given that, at the same time, all kinds of other messages, all kinds of other impulses are arriving via other dendrites? The more chemical the synapse, the more the brain ceases to be a determinate system and becomes a probabilistic one. An American scientist entitled one of his articles – as usual, you’ll have to pardon my English – “An Uncertain System”, a system of uncertainty, a probabilistic system. Let’s say, for the moment… we’ll see that it’s not precisely this, but we’re making slow progress. A probabilistic system, fine.[12]
From the moment there is a critical gap to cross, we’re forced to switch to a kind of aleatory mechanism, which is the mechanism of the chemical mediator and the number of quanta it will release. Given that the brain is besieged by messages coming from all these dendrites, that it’s perpetually… each time there is a cut in the form of these irrational cut-points, you arrive at the question of how the linkages are made. How are neurons linked together? The answer is that linkages are no longer determined, as they were in the electrical mode. The linkages become probabilistic. But what is a probabilistic linkage? What do we mean by a probabilistic linkage?
For the moment, let’s just retain two conclusions: in what form do the two points of mutation in the cerebral model take place? The brain implies, or the brain fosters – yes, that’s even better – it fosters a space, or no, it develops, yes, develops, no… well, whatever, any word – develops a topological space and envelops a probabilistic space. So, in place of the vertical axis of integration-differentiation, we substitute the topological contact of outside and inside. For the horizontal sensory-motor axis, we substitute the probabilistic character of linkages since linkages are interrupted by irrational cut-points. In these two ways, we can no longer – if we think about life – we can no longer experience our brain in the manner of a tree, we no longer have a tree in our head: that’s all over. It’s finished.
A brain specialist… you’ll find a lot of information on synapses, for example, in Jean-Pierre Changeux’s well-known book, L’homme neuronal [1983], and you’ll also find information focusing more on the probabilistic aspect of the brain in the work of an American neurobiologist called Steven Rose, R-O-S-E like a rose, whose book The Conscious Brain was translated into French a few years ago by Editions du Seuil.[13] And in Steven Rose’s book, there’s a very interesting metaphor. He says: You understand, there are very special cases where an axon… because in each case he tries to assess the probability of the message or impulse passing. He says that this varies according to several factors. There are, for example, cases where the dendrite of neuron B… the dendrite of neuron B winds… winds around the axon. Can you see it? It coils. Now, dendrites all have spines. So, on each spine of the dendrite, there’s a synaptic junction, there’s a synapse.
So, in the case of a single dendrite and axon, you can have a multiplicity of synapses, when the dendrite coils with its spines. And then the metaphor – today what I’m saying may sound rather dull, but at the same time it’s always a source of small joys – where Steven Rose says something quite wonderful. He says: “It’s just like when”– wait… I have to remember the quote… – “twisting around each other like bindweed around brambles”.[14] That’s beautiful! And it’s going to come in handy, because it’s not a metaphor. The bindweed, you see, is a weed. The bindweed wraps itself around the bramble, and the bramble is exactly the dendrite with its spines. If I drew you a picture of a dendrite… but you can imagine it for yourselves, an extension with spines. This particularly bizarre dendrite wraps itself around the axon of the preceding neuron, and at each point of spinal contact, there is a synapse. So, the dendrite is the bramble, and the axon that coils, is like the bindweed that coils around the bramble. How far we are from the tree model! We’re in a completely different field.
In other words, feel, feel the world of the future, it’s not a tree we have in our head, what we have in our head is grass! And this is the way we must live, through a lived relationship with the brain. When I say “lived relationship”, I’m thinking of a great author, Henry Miller. He had a prodigious gift: he always lived his brain as though it were grass. Very interesting, though it’s not just an image. He lives as though the inside of his head was filled with grass. Americans are very good at this. Americans, it’s well known, don’t believe in trees, they believe in the prairie so… Well, they’ve always experienced what’s going on in their heads as grass. Ever since one of the greatest poets… since the… since America’s founding poet, that’s the way it has been.[15]
It’s very odd. And when… and when Miller asked himself in what form he would be reborn, because he pretended to believe or he pretended to believe in rebirths, he said it would be “as a park”, he would be reborn in the form of a park.[16] Some people want to be reborn as a blade of grass, and some people want to be reborn as a tree, that’s their business. But then, they can’t have the same synapses, can they? Because if we function through chemical synapses, then the tree-brain is finished. The most we can hope for is to witness, even encourage, the marriage of a bramble and a bindweed. And what could be more beautiful? So, either you prefer the strength of a poplar and you’ll be reborn as a poplar, which you already are, or you love the marriage of bindweed and bramble. The choice is yours! But in each case, you will think in a completely different way. Though there will still be opportunities to understand each other! Okay.
So, here are my provisional conclusions. Once again, we find ourselves faced with a completely different schema, a brain model where once again, the horizontal axis… the horizontal axis has become probabilistic and the vertical axis has become topological. From this point, it’s no longer a question of a mechanism of integration-differentiation. It’s a question of outside and inside. Through the relative distribution of the internal and external milieus. And in the other case, it’s no longer a question of sensory-motor linkages, so what will it be? It’s a question of probabilistic communication, and here we’re faced with a problem – before we have a short break – well, how does this kind of probabilistic communication come about and how can it be conceived? What is this mode of linkage that is no longer a sensory-motor type of linkage? That’s the point we’re at now. How about a little pause? Not for long, though. Make sure it’s no more than six minutes, alright? Six minutes… [Recording interrupted; 1:41:43]
… the formula, looking for a formula concerning a linkage that would no longer be of the sensory-motor type. So, you see, this is consistent with all that preceded, since all that preceded was based on a rupture, and on the existence of a sensory-motor rupture. So, in the case of a sensory-motor rupture, how can we conceive of linkages, and in what form? Unless, of course, there are no longer any linkages at all. But now I’d like to multiply the paths for research by suggesting four or five very different directions.
First direction: we see bizarre things going on in fields… for the moment, let’s call them “intermediate fields”. But between what are they intermediate? Between chance and determination, chance and dependence. And what do we mean by these terms? An example of chance: a lottery with successive chance draws, these draws being independent of one another. The very condition is that the draws are independent. The same applies to the game of roulette, where the throws are independent of one another. We can always derive… from such a field of chance we can always derive what we call laws of probability. But I’m mixing everything up. These so-called laws of probability are well known and subject to certain conditions: the independence of the particles or the throws, the fluidity of the surrounding space, the speed of diffusion and so on, and its connection – which we know well from the kinetic theory of gases – with the idea of Brownian motion, Brownian motion of particles in a fluid state. Then, on the other hand, we have determined linkages.
So, what is there in between? A whole field was identified, but it had yet to be recognized, under the name of a field of partially dependent aleatory phenomena, or semi-aleatory phenomena, partially dependent aleatory phenomena, that is, phenomena that don’t lead back… that lead back neither to the semi-aleatory phenomenon… mixtures of dependence and chance. First path: how can we obtain such a mix of the aleatory and the dependent? By operating through random draws, but successive random draws that depend on one another. Here we are in the domain of mathematics. What happens in the case of successive random draws that depend on one another? This is what we call “semi-aleatory phenomena”, aleatory phenomena that are partially dependent… [Recording interrupted; 1:47:02]
Part 3
… So, in a lottery, you make a draw, a first draw, A, a second draw, B, a third draw, C. Each of these draws is strictly independent of the others. So that, after a while, you have to change the person drawing so that there’s no muscular dependency on their part. Imagine the other situation. A strange chain… it will form a chain. A chain of random draws, but each random draw depending on the one before it. Such a chain… it has become well known in mathematics and in cybernetics under the name of one of its inventors: [Andrei] Markov, M-A-R-K-O two Fs or a single V – it’s up to you – a Markov chain. And to help you understand this, I’ll give you a quick example.
Markov began by studying an odd question: how would it be possible to create a fake Latin, or fake English, or fake French? How can you make a fake language? It’s well known as the “mumbo-jumbo problem”. How do you make mumbo-jumbo? You can always conceive… so you give yourself a series of urns there, urns or boxes. I suppose I give myself the following rule: each box will contain groups of three letters, the first two of which are constant… the first two of which are constant. So, in your first urn, you put a-b-a, a-b-b, a-b-c, a-b-d, a-b-e, a-b-f and so on. You see? In the second one, you put b-c-a, b-c-b, b-c-c, b-c-d, and so on. So each urn contains a group of… let’s call them “trigrams”, a group of three letters, the first two of which are constant. You follow me?
If you only do that, it won’t yet give you anything. But as you know, regarding languages, this method is capable of calculating the frequency, for example, in terms of letter groups. In French, we would say that the letter Q, in a maximum number of cases, is followed by U, and we’re able to calculate the frequency of the Q-U group in relation to groups including Q that would not be followed by U. Right now, the only one I can think of is COQ [rooster]. Similarly, in French, there’s a significant frequency of the letter H being preceded by C. You can calculate – and this, of course, is where computers are useful which is why all this was very much linked to computing problems, and Markov chains would never have been discovered before the advent of computers – you can establish the frequency of C-H. So, let’s say you make frequency calculations with a language, it’s been done a lot. You can also make frequency calculations regarding the language of a particular author. It’s been done for [James] Joyce, under very interesting conditions. You can make frequency calculations not just on groups of letters, but on words. This has been done for [Stéphane] Mallarmé, for example. It’s not clear where all this is leading… Yes, it will allow you to make mumbo-jumbo, fake Mallarmé, fake Joyce.
But how do you go about it? You take a frequency threshold which ensures that your trigram, that is your group of three letters, has a certain frequency in the language in question. Then you go ahead and draw from an urn, you take an urn, you take the urn… and each urn is defined by the first two letters of the trigrams. You take the letter, you start with the I-B letter… no, I mean with the I-B urn. In the example I have here, taken from a book by [Raymond] Ruyer, which I’ll tell you about… [Recording interrupted; 1:53:22]
… and then, in this case, we draw U. So we write IBU. The first draw gave you IBU. This is a lottery draw. IBU, I-B-U. Then you have two options: either you can take any urn and draw again, at which point it will be a succession of independent random draws; or – I hope you’ve already guessed – you can take the B-U urn and draw from that. Suppose you take out BUS. From the I-B urn, you’ve drawn IBU; from the BU urn, B-U, the last two letters, you’ve drawn BUS. Now you have to take the US box, U-S, from which you’ll take out U-S-C, okay? U-S-C. Then from the S-C box, you’ll take out S-C-E. Okay, so now you have a chain. Here you can obviously claim the right the right to cut it. This gives you, I’m reading slowly–I’m appealing particularly to those who have studied some Latin – IBUS, I-B-U-S, cut, you cut there; CENT, C-E-N-T, cut; you’re going to continue your draws, IPITIA, I-P-I-T-I-A, cut; VETIS, V-E-T-I-S, cut, and so on.
As we say, it looks unmistakably Latin, and you can see why, it’s not complicated, it would be amazing, since you’ve retained trigrams based on frequencies, on statistical frequencies carried out across the whole language. You can do the same with fake English, it’s what we’ll call gibberish. You’ve done Latin gibberish. In the same way, you can make Joyceian gibberish. Even this Latin gibberish, in this example, a particularly elaborate example developed by… in a book by a linguist called [Pierre] Guiraud, Les caractères statistiques du vocabulaire.[17] Well, you can make different kinds of gibberish. What is this all about? Why am I telling you all this? I’m telling you this because here we have a typical case that should help us understand what we mean by a “Markov chain’, better than if I defined it by mathematical formulas. You see how each draw is made at random, from the urn, but at the same time each draw depends on the previous draw, since for the second urn you have chosen the one determined by the last two letters of the trigram chosen in the first urn.
This is a clear example of what we will call semi-dependent aleatory phenomena, or partially dependent aleatory phenomena. Each time there is a random draw, but under conditions of dependence between the urns and the successive draws. Here we’re no longer in a lottery, nor in conditions of pure determinism. It’s semi-aleatory, a Markov chain. You could say that it’s literally a mixture, a structured mixture. We can say it’s a structured mixture in the sense that it’s not structured as a language, and yet nor is it a purely random mixture.
Second path for research, just to be quick. At first glance, this seems to be unrelated. In the field of cosmobiology, regarding the eternal problem of how… how a living organism comes into being, biologists have long been making – and also physicists and chemists – for a long time they have been making a very interesting hypothesis. Long before Markov chains, they too made use of a linguistic comparison. They said, if you give yourself a random bunch of letters from which you then draw, you have very little chance of coming up with a word. But if instead you give yourself syllables, you obviously have a much better chance of forming a word. And this gave them the idea of an “intermediate state”. In other words, they said, wouldn’t it be possible to conceive of the emergence of living things in the following way: through a passage through intermediate states, provided these states are stable, and we move from one intermediate state to another by means of fluctuations, by means of fluctuations around the mean of the previous state?
What does this mean in terms of cosmobiology? It means that, for example, we start with a particular state of body that we call “colloids”, which contain nothing of life, colloids, that would be the first state. Under certain conditions yet to be determined, there is a fluctuation, what we’ll call a fluctuation in relation to the mean state of the colloid. And this fluctuation gives what we call a “coacervate”, for example, a coacervate of compounds, carbon compounds. Now, fluctuations in relation to a mean state are a constant occurrence – I mention this right now because it’s essential – they occur constantly. Even in a gas, with a Brownian particle distribution, with random particle distribution, you always find fluctuations. What do we call a fluctuation? A moment when an unusual number of particles group together in a precise region. But these fluctuations are usually immediately neutralized or compensated for. This is what prevents the intermediate state from taking hold. As soon as it has formed, it has already dissipated.
So, the problem of cosmobiology will be: Can we imagine conditions where a fluctuation forms in relation to a given state, and the fluctuation no longer dissipates but is amplified in such a way as to give rise to a new state? This is a fundamental question. So let’s leave aside for the moment the problem of conditions, since we’ll be coming back to it. Under what conditions does a fluctuation, instead of being cancelled, become amplified? Let’s try to understand it better. We start with the colloidal state, fluctuating in relation to its mean state, which gives us a new state, a coacervate of carbon compounds, a set of carbon compounds. Once again, there’s every chance that the fluctuation will be cancelled. Let’s suppose that under certain conditions it isn’t cancelled. At that point, well, we will have intermediates, there will be new intermediates – I’m jumping ahead, to speed things up a bit – we’ll end up with the possibility of the formation of enzyme chains, or protein chains, protein chains that, however, won’t yet form a living organism.
But from intermediate state to intermediate state, in other words from amplified fluctuation to amplified fluctuation, can’t we conceive of the problem of the origin of life in another way, a new way? You see how this is very different from chance formation. So, what is it? What I’m describing to you is a Markov chain, but here it’s in terms of evolution, it’s a Markov chain. You have exactly one series of mutually dependent draws, a series of mutually dependent random draws, where each time the fluctuation marks a chance which, providing it isn’t cancelled out, if conditions are such that they permit amplification, will lead to a new draw which will once again produce another… you have a sequence of draws that depend on one another, even though each individual draw is random. You have a Markov chain. Or some kind of Markov chain. For the moment, we’ll keep… here, everything I’m saying will be worthless if we forget our problem: that’s all very well, but for a fluctuation to generate a new state, which will be in semi-aleatory relation, you see, it will be precisely in an aleatory relation that is partially dependent on the previous state. The coacervate of carbonaceous compounds will be in semi-aleatory, semi-dependent relation with the colloidal state, provided that – again, I haven’t really answered the question yet – the fluctuation isn’t cancelled out.
There’s a text by [Charles] Darwin, a letter by Darwin, which I think is brilliant, where Darwin as a young, actually not so young, man recounts how he is constantly being asked: If life had simply begun, then why couldn’t it begin again today? It was the anti-evolutionists of Darwin’s time who were already resorting to this argument: If it happened once, why shouldn’t it happen again? And Darwin replied how it would be very difficult today for another form of life to emerge. And why is that? Suppose that in a small pond – this text is brilliant, because it gives us the answer – suppose that in a small pond – obviously the same thing wouldn’t happen in the open sea – suppose that in a small pond, coacervates form from colloidal states. Well, today, they would have no chance of producing an amplifying fluctuation. Why not? Because they’d immediately be eaten, they’d immediately be eaten by the living creatures that were already there. They would be immediately absorbed, there would always be a fish to swallow them. So there’s a simple reason why life has only been able to form once: once it is produced, it consumes everything. So, there’s not much chance of another life form with chains of a different type emerging. The chains of a different type would immediately be swallowed up. But his remark, “in a small pond”, should once again enchant us, because it sets us on the path towards the solution. Under what conditions can a fluctuation become amplifying? But this is just a path. We don’t know yet, we simply don’t know yet.
So that would be the second research path: cosmobiology. Now I conclude this point here, once again, because I aim to go very slowly. You can see that this linkage of intermediate stable states, where we go from one to the other by means of amplified or amplifying fluctuations, form a Markov chain, or a kind of Markov chain, in other words a sequence of partially dependent aleatory phenomena.
The third type of research is that undertaken by [Ilya] Prigogine and Isabelle Stengers. This research focuses precisely on fluctuations – it’s in chapter 6 of La nouvelle alliance[18] – where they directly pose the question: Under what conditions does a fluctuation become amplifying, and under what conditions, on the contrary, is it cancelled or balanced out? So, here they should be able to give us an answer to the question we posed earlier. The state, the primary state where fluctuations are immediately balanced out is once again the state of the kinetic theory of gases. A large number of particles distributed in a fluid medium, fluid – ultimately, we could say ideally fluid – a large number of particles in an ideally fluid medium, what happens to their motion? Each particle follows what we can call free motion. What is free motion? It’s the average distance travelled by a particle before another particle collides with it.
You see why I insist on this free motion. Why do we need the fluid medium of a gas? Because in a solid, particles are no longer independent. Why are particles no longer independent in a solid, or even in a liquid? Because the actual state of the solid or liquid brings them together in a region where they lose their very independence, they are caught in the region of the solid in such a way that they can no longer follow free motion, they lose their free motion. In a gas, with Brownian motion of the particles… You see, Brownian motion is therefore when a particle follows a trajectory until it is, until it collides with another particle that sends it off in another direction and so on. And you have the type of random distribution subject to the laws of probability insofar as you have a very large number of particles. A fluctuation occurs when a large or at least a significant proportion of this large number of particles find themselves in the same region.
I would say that here it’s immediately corrected, it’s cancelled out. Why? Because, Prigogine and Stengers tell us, because of the effect of diffusion. Indeed, in a fluid medium, it’s because of the diffusion that binds all the regions of the system. It’s because of the diffusion that binds all the regions of the system with the environment, in particular the fluctuating region, which is to say the region where the abnormal concentration of particles has appeared. And it’s because there’s such a high level of diffusion between the region where the fluctuation takes place and the environment that the fluctuation is immediately balanced out and the probabilistic distribution of particles re-established. So, it’s a question of diffusion.
So, the answer to my question is that a threshold has to be crossed for a fluctuation not to be cancelled or balanced out but to be amplified. What would it take for a fluctuation to be amplified to the point of generating a new state? The fluctuating region would have to be somehow protected from the rapid diffusion that re-establishes the mean state. Prigogine goes even further, because he says: You know, this applies to our societies too. How can we explain the fact that our societies, which are very complex, are so stable, when at every moment there are fluctuations, that is, deviations from the mean? Well, he says, there’s an equivalent to what happens in fluidity: it’s the speed of information. That’s why the speed of information is fundamentally something… it’s linked to maintaining order. It’s the speed of information that immediately cancels out the fluctuations and returns them to…. that crushes them.
And then, in an interesting sentence, he says, Well yes, this is why the fluctuations that are likely to survive very often have their origins in marginal groups, because marginal groups are protected against the speed of information, of diffusion. So it will spread afterwards, but precisely, it will then be something amplified [un truc amplifié], or – what was the term again – an amplifying or amplified fluctuation. Do you understand what I’m saying? So here we have the answer to our question: for a fluctuation, meaning a deviation from a mean state a to be amplified, it must somehow escape the speed of diffusion. Under what conditions can it escape the speed of diffusion? It’s that, if there’s an accumulation – and here we have the answer, I can’t think of any other – if there is an accumulation in a restricted milieu and this restricted milieu is protected from the surrounding milieu that would cancel out the fluctuation or compensate for the fluctuation, which is to say ensure its cancellation by rediffusion. So the restricted milieu has to be protected from diffusion.
So there it is again in full force – I’m surprised he doesn’t refer to it directly, I don’t think he does, even in his other… – you have a Markov chain in full effect. Prigogine gives a very curious example, he gives the example of termite mounds, which is typically…. which can be interpreted in terms of a Markov chain. It’s very interesting, because you might think that the termite mound is a classic example of a centered system, an arborescent, centered system. No, not necessarily. There’s an interpretation of the termite mound in… which sees it as a typical Markov chain. That is, termites transport little balls of earth that they then drop along their path. Do you follow me? Imagine you’re a termite, you transport balls of earth, you drop them along the way. And as they drop them, the termites mark then with a hormone. This hormone has the property of attracting other termites who drop their own. So, what does it take for the pillars, at least the pillars of the termite mound, to be established? All it takes is an aleatory factor, a fluctuation, meaning that by chance, by pure randomness, a large number of termites drop their balls of dirt in a restricted portion of their territory. This marks a fluctuation in relation to the mean distribution of earthballs. Do you follow me? We’ve seen a lot of termites today, haven’t we?
There we have the random factor, that a large number… that a considerable number of termites each drop their ball of dirt in a restricted portion of the termites’ territory. From that, everything else links up, everything links up, meaning that, if this happens, you have a portion of the territory that by its very nature already resists diffusion. Why does it resist diffusion? Because of the hormone, which attracts other termites to drop their ball of earth too. And then you’ll have equally distanced pillars rising up. Why equal distanced? The distance will be equal to the olfactory range of the hormone. The beginning, at least – because you have to, you have to go very slowly, you have to be careful – the beginning of the construction of a termite mound can be likened here to a Markov chain, that is, a succession of semi-dependent aleatory phenomena. As you can see, the determining factor was the hormone, the aleatory factor was the random release of earth balls. And the condition for the fluctuation to be amplified was that a certain number of termites released their ball of earth in a very restricted, very… That’s the third path.
Fourth path of research. This is the research of another cyberneticist, a cybernetic biologist by the name of Pierre Vendryes, V-E-N-D-R-Y-E-S… He shouldn’t be confused with another Vendryes, who was his father and he was a linguist. Vendryes has a very simple schema. I’m telling you this because it’s quite beautiful, and at the same time, it helps us move forward. He says: Let’s try to define living beings by the constitution of internal milieus. The organism is an instance that has constituted an internal milieu, and the more solid the internal milieu – and we see what he means by this, it’s right at the heart of our problem… you see, this takes us back to the notion of internal milieu, but at another level – the internal milieu is now in the territory of a living being, it forms part of the territory of the living being, and it becomes all the more consistent the more it acquires self-regulation, so it escapes the fluidity of the external milieu, or it escapes diffusion. The constitution of an interior milieu is, literally – it’s strange that I keep forgetting the word… – an amplifying fluctuation.
What is the immediate consequence of the internal milieu? The immediate consequence of the internal milieu is that the living being will enter, or will be able to enter, into an aleatory relationship with the external milieu. A living being with its own internal milieu thereby enters into an aleatory relationship with the external milieu, rather than a deterministic one. Why is this so? It depends on the point of view from which you look at it. An internal milieu implies the constitution of reserves. The living being therefore has reserves. As long as it has reserves, it doesn’t require an external life. So, you see, the conditions are very precise.
Student: Inaudible remark.
Deleuze: Thank you. You see… I don’t know what you see anymore.
Yes, of course, if the living being is hungry, for example, it is clearly not in an aleatory relationship with the external milieu. It’s hungry, so it seeks food outside. But that’s not currently our problem. We’re not considering the living being when it is hungry. We’re looking at living beings insofar as they have reserves, constituted by their internal milieus. So, at the moment and insofar as it has reserves constituted by its internal milieu, it is independent of the external milieu, as long as the reserves last. It will again become dependent once its reserves are exhausted. For example, if I have sugars, proteins and so on, in my body, I can wait to eat. I am independent of my external milieu.
So, what do I do? It’s common knowledge: I wander. I go for a stroll [balade]. What is a stroll? It’s entering into an aleatory relationship with the external milieu. If I’m hungry, if my reserves are exhausted, I stop walking. In the end, I either walk or I sleep. And if I don’t sleep, well, I just wander around. It’s like flânerie. Lions sometimes sleep, sometimes prowl, sometimes wander. What’s the result? In other words, I’m in an aleatory relationship with the external milieu. Is this… so what will this aleatory relation of the living being give rise to? The autonomy of the living being is defined by its aleatory relation with the external milieu. Well, Vendryes goes much further than this. He says that this is perfect, there is organic autonomy, which we’ve just defined, but there is also intellectual autonomy. This is verified by our hypothesis, because our hypothesis is that there is an aleatory relationship between an idea and the external world. This notion of double autonomy is a good one. It concerns the brain, the brain as an internal milieu that enters into an aleatory relationship with the external world. This aleatory relationship is effectuated by what we call hypotheses.
So, what will the pace of this strolling movement resemble? Very strangely, it will resemble Brownian motion, but it won’t be Brownian motion. You see? And Vendryes, in order to define this type of motion… it won’t be Brownian motion because Brownian motion is defined by the independence of particles in a medium of diffusion, in a fluid, or ideally fluid, medium. What we have here, on the contrary, is the aleatory relation of a particle with a counter-aleatory internal milieu. For the internal milieu is indeed counter-aleatory. Do you follow me? I don’t know, perhaps you’re already too tired, but I’ve almost reached the end.
Vendryes was finally… yes, he says, look, and he took as an example…. he was one of the first to photograph flies or mosquitoes in flight, the flight patterns of flies and mosquitoes, and clearly here we have a movement that closely resembles, or that seems to closely resemble, Brownian motion. You see? Under ideal conditions of fluidity, flies and mosquitoes seem to behave very much in the manner of Brownian motion. You’ve already seen the way flies fly. There’s an absolute difference, isn’t there? Again, we have an absolute difference, since instead of independent particles, we have a double movement, an aleatory relation with the external milieu insofar as it depends on a counter-aleatory internal milieu. And he will call this Brownoid motion, going so far as to say that history, the history of mankind, is full of Brownoid movements.
And in a rather bizarre book, De la probabilité en histoire[19] [On Probability in History], he analyzed Bonaparte’s expedition to Egypt, the Egyptian Expedition and Nelson’s famous pursuit of Napoleon in Brownoid terms, saying… saying, in fact, that it was clearly a chain of Brownoid… quasi-Brownian paths that Napoleon took and that Nelson took in pursuing him, each unaware of where the other was, and each traversing the Mediterranean according to chance distributions that were determined by the reserves they had. Well, exactly. This is something you can find in history.
To speed things up, what would this imply? I find all this very interesting. It means that history never ceases being traversed by semi-aleatory mechanisms. Or, if you prefer, [Raymond] Ruyer has a lovely phrase in his book La genèse des formes vivantes [20] [The Genesis of Living Forms]: “History is not logos, it’s gibberish.” History is gibberish, yes, it’s all gibberish, it operates. It’s neither chance nor determinism, nor is it the idea, meaning it’s not logos. It consists of partially dependent aleatory phenomena, of Markov chains.
Regarding what Vendryes, still imperfectly, it seems to me, referred to as Brownoid motion, I would say that it’s in our interest to systematically theorize it in the form of Markov chains. The relation with the milieu, the aleatory relation with the milieu, will be of the Markov chain type, that is, it will be a partially dependent aleatory phenomenon. A perfect example of a partially dependent aleatory phenomenon would be the daily itinerary of a taxi. The trajectory a taxi takes in a single day is very close to Brownian motion. Sociologists are fascinated by this and many people have studied the cab’s daily journey. But as you can see, it’s not exactly Brownian, for a very simple reason. It’s much more a chain of partially dependent aleatory phenomena, since each journey depends in part on the preceding one, if only from the point of view of the pick-up point. As a result, a cab’s daily journey is much closer to a Markov chain than to Brownian motion. And this is what Vendryes was trying to explain when he spoke of Brownoid rather than Brownian motion.
So then, what does this all boil down to? Well, in a way, maybe we’ve got our solution. I mean, where does all this lead us? If I had to try and give a formula for Markov chains, not a scientific one, but a philosophical one, I’d say it’s a set of re-linkages with no prior linkage. And this is what is important to me. The rest is secondary, it was all a question of getting to this point. And this is what I find so mysterious: how is it possible for there to be re-linkages if there are no linkages? We must suppose that the linkages are random but the re-linkages are not. I’d say that this is what a Markov chain is. There is no longer any linkage, there are only re-linkages, by which I mean that notions independent of one another are re-linked, instead of there being a linkage of dependent notions. Instead of the linkage of dependent notions, there is a re-linkage of independent notions. Not for the first time, relinkage takes precedence over linkage, and Markov chains give us the mathematical structure of such an event.
So this is very useful to me, because it provides an answer to the question of how transmission would take place in the brain. Transmission in the brain is effectuated by relinkage in independent series. This would be much closer to Markov chains – hence the idea of a probabilistic brain, much closer to a Markov chain than to electrical transmission. So now we have everything. Because now we will pass to the question I briefly summarized: What does all this mean for cinema? Couldn’t we also say that there are two very different image regimes from this point of view? There was a regime of image linkage that inspired the classical conception of montage, and this was a sensory-motor linkage of images. But wouldn’t so-called modern cinema propose very different structures, which would be the re-linkage of independent images? There would no longer be linkage, only re-linkages. [End of the session; 2:38:18]
Notes
[1] Deleuze apparently had to cancel the December 4 session due to illness.
[2] On these models, see The Time Image, pp. 210-212.
[3] See Roman Jakobson, Essais de linguistique générale, Paris: Minuit, 1963 Tome 1, 1973 Tome 2.
[4] See Simondon, L’individu et sa genèse physico-biologique, Paris: PUF, 1964 / Grenoble: Millon, 1994.
[5] Ibid., in the Millon Edition see p. 224.
[6] On this subject, see Simondon, op. cit., in the Millon Edition, see p. 223. See also The Time-Image, p.318, note 31.
[7] See Simondon, Millon Edition, p. 225.
[8] Ibid., in the Millon Edition, see p. 224.
[9] Ibid. In the Millon Edition, the section “Topologie et ontogenèse” corresponds to pages 222-227.
[10] The author Deleuze refers to here is Steven Rose. The notion appears in his book, The Conscious Brain, which Deleuze will cite later in the lesson.
[11] On this discussion, see The Time-Image, pp. 211-213, in particular p. 318, note 32.
[12] Deleuze cites the expression “uncertain system” in The Time-Image, p. 318, note 32, without providing a source except Jean-Pierre Changeux, L’homme neuronal, Paris: Fayard, 1983.
[13] Steven Rose, The Conscious Brain, New York: Knopf, 1973. Steven Rose is actually English.
[14] Ibid. p. 76
[15] No doubt Walt Whitman and his Leaves of Grass.
[16] See H. Miller, Tropic of Capricorn, London: Calder, 1964, p.111.
[17] Paris: PUF, 1954
[18] Translated as Order out of Chaos: Man’s New Dialogue with Nature, Toronto/New York: Bantam Books, 1979.
[19] Paris: Albin Michel, 1952.
[20] Paris: Flammarion, 1958.
French Transcript
Gilles Deleuze
Sur Cinéma et Pensée, 1984-1985
6ème séance, 11 decembre 1984 (cours 72)
Transcription : La voix de Deleuze, Laura Cecilia Nicolas (1ère partie), Guadalupe Deza; correction Anselme Chapoy-Favier (2ème partie), Sabine Mazé (3ème partie) ; révisions supplémentaires à la transcription et l’horodatage, Charles J. Stivale
Partie 1
… ne pas vous déranger, mais c’est la plus belle forme de la conscience morale, la politesse, hein ? Sinon ce n’est pas que je crois que… [Pause]
Alors on a perdu du temps [À cause d’une maladie, Deleuze a dû apparemment annuler la séance du 4 décembre], mais on va le rattraper parce qu’on va faire aujourd’hui une séance très vague, puisque je ne suis pas, je ne suis pas guéri. Et cette séance très vague donc, elle porte — et puis j’ai une idée que je vous expliquerai à la fin de cette séance — mais cette séance-là aujourd’hui — qu’on va faire très douce, hein, il ne faudra pas me demander beaucoup — c’est sur notre dernier point, le dernier point de ce programme. Et je voudrais qu’on le fasse aujourd’hui, donc j’irai plus vite, pas plus vite, pas plus vite, mais j’irai plus vague, quoi, pour regagner le temps. Ce dernier point qui nous restait, c’est le quatrième point de mutation [1 :00] concernant le cerveau. [Pause] Alors, on aura l’occasion de récapituler tout ce qu’on a fait ce trimestre, mais je ne récapitule pas encore, je suppose que vous vous rappelez, c’était notre quatrième point, le dernier pour nous.
Qu’est-ce qui s’est passé ? « Mutation » n’est peut-être pas même un mot heureux. Ça s’est fait d’une manière tellement éparpillée, et puis insensible, tout ça, mais ce qui m’intéresse, vous le savez, parce que je l’ai dit, c’est trois points. Qu’est-ce qui a changé du point de vue, d’un triple point de vue ? Du point de vue d’une biologie du cerveau. Ça, c’est les savants, c’est l’affaire des savants, mais enfin, je n’ai jamais vu de cas où les problèmes fussent exclusivement [2 :00] scientifiques. D’autre part, donc, pas seulement du point de vue d’une biologie du cerveau, mais du point de vue d’un rapport vécu avec le cerveau. [Pause] Et enfin, du point de vue d’un cinéma comme cerveau ou d’un cerveau comme cinéma. Je ne dis pas que tout cinéma soit cerveau et tout cerveau soit cinéma. Mais, si vous vous rappelez ce qu’on a vu, on a vu certaines choses, par exemple, un cinéma qu’on pouvait appeler très rudimentairement « un cinéma des corps ». Et je disais, de même qu’il y a un cinéma des corps, il y a aussi un cinéma du cerveau, bon.
Là, mes trois points c’est donc jouer de ces trois dimensions : les changements éventuels, les grands changements éventuels dans une biologie du cerveau, [3 :00] les grands changements éventuels dans notre rapport vécu avec le cerveau s’il y a lieu, et les changements éventuels dans un cinéma du cerveau. Et pour ça, comme dans les autres cas, j’essaie d’opposer, toujours très grossièrement, une image dite classique, en laissant à « classique » tout son vague, et une image dite moderne en laissant à « moderne » tout son vague. Ce qui sous-entend que, par exemple, s’il y a un cinéma moderne du cerveau, il ne procède pas de la même manière que ce qu’on pourrait appeler le cinéma du cerveau d’avant-guerre. De même que les modèles de cerveau ont sérieusement évolué dans les sciences, ça tout le monde sait. Mais ce que tout le monde peut sentir, même sans le savoir, c’est que notre rapport vécu avec le cerveau a changé aussi, [4 :00] enfin, dans certains cas. [Pause]
Et je me dis, quelle était l’image classique du cerveau ? Comment est-ce qu’on pourrait la présenter ? Comment est-ce qu’on pourrait la représenter, ben, immédiatement du point de vue de la science ? Et, mon Dieu, je ne vais pas dire des choses compliquées là-dessus. [Pause] Je dis, dans l’image scientifique du cerveau, dans l’image classique, il me semble qu’il y a deux axes. L’un correspond à l’action des centres, l’autre correspond à l’action [5 :00] des éléments. Et ce sera déjà tout un problème dans la neurobiologie que de savoir en quoi consistent les centres, en quoi consistent les éléments, quelles sont leurs différences, [Pause] quels sont leurs rapports. Je me contente de dire : les centres qui définissent des aires cérébrales, a-i-r-e, qui définissent des régions, des aires cérébrales, [Pause] se présentent sous un double aspect : d’une part, et un aspect indissociable. Ils sont « intégrateurs », ce sont les fameuses [6 :00] intégrations cérébrales. Mais ils sont aussi différenciateurs, ils différencient des aires. Par exemple, on différenciera une aire [Pause] projective, c’est-à-dire concernant la projection des organes des sens, [Pause] une aire motrice, une aire associative. [Pause] Mais l’action d’intégration donc, elle est strictement indissociable d’une [7 :00] action de différenciation. Voyez d’où, dans le schéma que j’ai tracé, ma flèche verticale [Deleuze se déplace vers le tableau], ma flèche verticale : intégration, différenciation, [Pause] comme fonctions des centres.
D’autre part, la flèche horizontale, elle concerne les éléments. Tout élément, tout élément nerveux, tout élément cérébral est à la fois sensoriel et moteur. Ce sont des éléments sensorimoteurs qui entrent dans des enchaînements [Pause] [8 :00] ou dans des associations : associations ou enchaînements sensorimoteurs, donc associations ou enchaînements sensorimoteurs qui concernent les éléments, intégration et différenciation qui concernent les centres. Vous devez me dire que ça ne vous étonne pas… [Interruption de l’enregistrement] [8 :27]
… car nous retrouvons, au niveau du cerveau, des données les plus simples d’une biologie du cerveau, ce qu’on a vu sur l’image classique du savoir. Tout se passe comme s’il y avait isomorphie entre le modèle du savoir et le modèle du cerveau. [Sur ces modèles, voir L’Image-Temps, pp. 273-276]
Car si vous vous rappelez, quand nous avons traité du modèle du savoir à propos d’une autre mutation, l’image classique du savoir [9 :00] nous a semblé consister en quoi ? Un axe, que j’appelais « l’axe du concept », d’après lequel s’opèrent les intégrations de concepts et les différenciations, les divisions de concepts, c’est-à-dire les spécifications, spécifications et intégration, qui exprimaient le mouvement du concept dans le savoir. Et d’autre part, un autre axe, qui se présentait comme les associations d’images. Comment se faisait le rapport des axes et la circulation à travers eux ? C’était très simple, et c’était le modèle du savoir. Un concept ne se différenciait pas, c’est-à-dire, [10 :00] ne se divisait pas, ne se spécifiait pas, sans en même temps s’extérioriser dans une suite d’images prolongeables. [Pause] Mais inversement, une suite d’images ne se prolongeait pas sans s’intégrer dans un concept. On avait donc toute une circulation, qui était la circulation du savoir.
Ben, quand je dis qu’il y a isomorphie avec le modèle cérébral, en effet, [Pause] les centres ne se différencient pas sans s’extérioriser dans les chaînes [11 :00] sensorimotrices prolongeables ; les chaînes sensorimotrices ne se prolongent pas sans s’intégrer dans les centres. Donc, c’est normal, ce serait inquiétant qu’il n’y ait plus cette isomorphie entre le vieux modèle du savoir et le modèle [Pause] du cerveau. Je dirais pour des raisons-là que je n’ai même pas envie de développer, je dirais que… non, là j’ajoute quelque chose mais il nous manque un point. Si vous m’avez suivi, si vous avez un peu le souvenir de ce qu’on a fait précédemment, il nous manque déjà un point. Alors, c’est un devoir, mais est-ce qu’on pourra le combler ? C’est qu’il faudrait [12 :00] encore que, dans ce vieux modèle du cerveau, les coupures intervenant dans les chaînes sensorimotrices en vertu de l’autre axe, de l’axe vertical, il faudrait que les coupures soient des points rationnels.
Et là, on aurait rempli l’image classique du cerveau qui consisterait en intégration-différenciation par les centres, [Pause] association sensorimotrice, coupure rationnelle. Coupure rationnelle, j’ai justifié les deux premiers points : coupures rationnelles, points rationnels ; je n’ai pas pu encore le justifier. Mais si je pouvais, alors on serait tous très contents. [13 :00] Alors je voulais que vous le sentiez, ce serait une grande joie, enfin, une petite joie. Mais au moins les deux premiers points, on les tient.
Je dis, ce n’est pas à cause des schémas, mais si j’essayais de résumer quels rapports vécus on a avec le cerveau, au niveau de cette image, c’est tout simple : c’est l’arbre du cerveau. Vous me direz, pourtant ça ne ressemble pas à un arbre. Ben si, ça ressemble à un arbre, on verra comment. C’est-à-dire, c’est une manière de vivre, on a un arbre dans la tête. Cet arbre, c’est cet axe marqué par des points rationnels, des coupures rationnelles. Et beaucoup de gens ont vécu avec l’idée et vivent encore avec l’idée que leur cerveau est un arbre, [14 :00] c’est-à-dire, un système centralisé, système centralisé ce qui signifie : centre d’intégrations et de différenciation, et associations sensorimotrice, l’un s’exprimant dans l’autre, et formant une circulation, toute une arborescence du cerveau, toute une arborescence du cerveau suivant ces deux axes.
Or, sans vouloir du tout analyser en détail, ça pose toutes sortes de problèmes. Je cite, par exemple : quel rapport y-a-t-il entre les centres et les éléments ? [Pause] En admettant que les éléments soient des cellules cérébrales, [15 :00] ou des cellules nerveuses, les centres, c’est quoi ? Est-ce que c’est des cellules particulières ? Ça va être… Vous sentez là qu’il y a tout un problème historique des localisations — que je ne prétends pas aborder — des localisations cérébrales. Mais le problème est très, très compliqué pour une raison très simple. Suivant la remarque d’un très grand neurologue, d’un neurologue psychiatre, qui avait beaucoup subi l’influence de Bergson et qui s’appelait [Constantin] Von Monakow, il a écrit un livre admirable qu’on ne lit plus, je crois, un très, très beau livre, M-O-N-A-K-O-W. Il disait : vous comprenez, le tort, c’est de chercher une localisation [16 :00] spatiale des centres. [Pause] Les cellules sensorimotrices, oui, elles ont des localisations spatiales. Mais les centres intégrateurs et différenciateurs, ils n’ont pas de localisation spatiale. Et il proposait l’idée de localisation chronogène, une localisation chronogène. Alors il donnait des schémas, des schémas très compliqués. Vous voyez pourquoi il était bergsonien, il jouait avec une espèce de durée bergsonienne inscrite dans le vivant et dans le cerveau, et la localisation chronogène, ça attestait à un des thèmes les plus complexes de Monakow.
Pour vous faire sentir là toutes les difficultés, là, que… je dis, s’était imposée, par exemple, une notion célèbre chez un [17 :00] grand neurologue qui s’appelait [John Hughlings] Jackson, l’idée que dès lors, les maladies du cerveau étaient des processus de désintégration, de dissolution. C’est-à-dire, que les centres étaient atteints et que, dès lors, les associations sensorimotrices se libéraient du rôle intégrateur des centres. C’est ce qu’il appelait « dissolution ». Et la dissolution, c’était l’inverse de l’évolution. L’évolution, elle allait des éléments aux centres. [Pause]
Qu’est-ce qui compliquait, qu’est-ce qui faisait que… [Pause] [18 :00] Jackson disait quelque chose de très curieux. Il disait : les centres, c’est le moins organisé. C’est-à-dire, les éléments sont beaucoup plus organisés que les centres, ce qui paraît bizarre. Les éléments, à la fois intégrateurs et différenciateurs, ils sont beaucoup moins organisés. C’est qu’il s’expliquait très bien, sur ce qu’il comprenait par « moins organisé ». Il voulait dire que leur organisation se poursuit toute la vie. C’était une manière de rejoindre la localisation chronogène de Monakow — pas de rejoindre, parce que c’est, c’est Jackson qui précède Monakow dans l’histoire — mais c’était une manière de pressentir l’idée d’une localisation chronogène. Les centres, ils ne cessaient pas de se monter et de s’organiser toute la vie. Donc ils étaient moins organisés [19 :00] que l’axe sensori-moteur concernant les éléments.
Et encore Jackson, dans sa théorie de l’évolution et de la dissolution, vous voyez, il pouvait dire : l’évolution va du plus organisé au moins organisé, Et la dissolution, inversement, puisque le plus organisé, c’est l’automatique, c’est des chaînes sensorimotrices automatiques. [Pause] Et la dissolution donc, elle allait du moins organisé au plus organisé, tout comme l’évolution allait du plus organisé au moins organisé. Ça n’empêche pas que dans sa conception, il considérait, il me semble, ou il privilégiait singulièrement le rôle intégrateur des centres. [Pause] [20 :00]
Seulement voilà, là où je dis que c’est très compliqué, c’est qu’à mon avis – enfin, « à mon avis » — il n’y a pas des phénomènes de désintégration sans des phénomènes corrélatifs de « dé-différenciation ». [Pause] Donc les schémas de dissolution se compliquent énormément. Il n’y a pas une action de désintégration qui ne soit pas accompagnée d’une dé-différenciation, c’est-à-dire d’une suppression de la différenciation. [Pause] Et alors, bien plus, peut-être que dans les phénomènes de dissolution, il y a des troubles de désintégration prévalente, et des troubles de dé-différenciation prévalente. [21 :00] Ce qui compliquera beaucoup le schéma. Si bien que les cellules sensorimotrices échappant aux centres, ne sont pas désintégrées sans être aussi dédifférenciées, c’est-à-dire, perdent leur différenciation au même temps qu’elles perdent leur intégration. D’où si je saute dans un problème très célèbre, qui a été déterminant dans les schémas du cerveau, l’aphasie et les types d’aphasie. L’aphasie, vous le savez, ce sont des troubles du langage avec lésions cérébrales. [Pause]
Or, [Pause] classiquement, [22 :00] la grande théorie de l’aphasie distinguait — ça allait très loin, mais ça se comprend avec un axe aussi, déjà aussi compliqué — la grande théorie de l’aphasie distinguait six types d’aphasie [Pause] dont malgré tout, on pouvait dégager deux grands pôles. Et les deux grands pôles, vous en trouverez une étude linguistique assez poussée chez [Roman] Jakobson. Ce qui est une manière de suggérer que Jakobson s’inscrit et garde d’une certaine manière, garde implicitement, puisqu’il ne se prétend pas biologiste, garde implicitement… [Interruption de l’enregistrement] [22 :57]
… mais [23 :00] ils se laissent réduire à deux pôles, deux pôles qu’il [Jakobson] étudie particulièrement dans les Essais linguistiques [Essais de linguistique générale (Paris : Minuit, 1963, tome 1 ; 1973, tome 2)]. Et ces deux pôles vont s’expliquer puisqu’on va voir comment ils tiennent compte de notre axe, plutôt de nos deux axes : un axe vertical et un axe horizontal. Les deux grands pôles, c’est, nous dit-il, les deux grands pôles de l’aphasie, c’est des troubles que l’on pourrait appeler tantôt « troubles de la similarité », tantôt « troubles de la contiguïté ». [Pause] La similarité, selon lui, est à la base [24 :00] de ce qu’il appelle « métaphore », la contiguïté est à la base de ce qu’il appelle « métonymie ». [Pause]
Et si je résume très brièvement ces deux troubles, le premier trouble, dit de similarité, se présente comment ? [Pause] Le sujet a gardé le pouvoir de combiner les éléments de langage. Mais qu’est-ce qu’il a perdu ? [25 :00] Il a perdu… Il a gardé donc le pouvoir de combinaison, mais il a perdu le pouvoir de sélection. [Pause] Il sait combiner une phrase, mais il ne sait plus sélectionner entre un élément et un autre. Ça revient à dire quoi, ça ? Dans une conversation, il répond très bien, il continue une conversation, ou bien si on lui présente un fragment de mot ou un fragment de phrase, il complète très bien. Donc il a gardé l’art de la combinaison. [26 :00] Il a une gêne : c’est évidemment pour commencer une phrase, le mot initiateur. Pourquoi ? Sentez. Parce que le mot initiateur, il faut le sélectionner, il faut le choisir. Là il est gêné, il est embêté. Mais une fois que c’est parti, c’est parti. Il est gêné une autre fois : c’est lorsqu’il a à sélectionner entre deux possibilités. Par exemple, il emploie le mot « célibataire ». Pour quelqu’un de normal, « célibataire » est substituable à [27 :00] « non-marié ». Il ne peut pas sélectionner, il ne peut pas faire la substitution. Si on lui dit, « qu’est-ce qu’un célibataire ? », il entre en détresse. Il sait très bien construire sa phrase avec « célibataire ». Il sait combiner, il ne sait plus sélectionner. Or, comme dira Jakobson, c’est les deux grands pôles du langage : combinaison et sélection.
Bien plus, il ne peut pas faire la sélection minimum du type : « A = A », si vous voulez, le minimum de substitution, la substitution identique. Car si on lui dit : « dis ‘non’ », [Pause] [28 :00] il répond : « non, je ne peux pas ». C’est une situation en apparence paradoxale, puisqu’il ne peut pas dire « non », mais il dit qu’il ne peut pas dire en disant « non ». Vous voyez que c’est en fait sur deux axes très différents du langage. Puisqu’on lui dit : « dis ‘non’ », c’est-à-dire : « répète », substitution du type A est A, et il répond : « non, je ne peux pas », « non, je ne peux pas dire ‘non’ ». Le second « non » qu’il emploie, le « non, je ne peux pas », est pris dans une combinaison, ça il peut, mais la sélection de « non » et la substitution minimum de « non » par « non », il ne peut pas. [29 :00] Voilà, c’est un type d’aphasie, bon, aphasie de similarité, trouble de similarité.
Dans le trouble de contiguïté, ça va être l’inverse. La similarité semble sauvée, mais la composition, elle, … la sélection, elle, semble être sauvée, mais la composition ne l’est pas du tout. Dans ce type d’aphasie, vous rencontrez ce que les linguistes appellent « l’agrammatisme », agrammatisme, c’est-à-dire, la dissolution de la grammaire. [30 :00] La phrase de ce type d’aphasique – vous voyez que c’est très différent de l’autre cas – la phrase de ce type d’aphasique finit par être une espèce de morcelage de mots. [Pause] Ce qui disparaît, ce sont les liens entre les mots, et notamment ce sont les flexions. Quand il emploie un verbe, ce type d’aphasique le met généralement, le mettrait très souvent à l’infinitif. [Pause] Bien. Cette fois, comme dit Jakobson, la similarité, elle est sauvée. Il peut très bien dire le mot « crayon », [31 :00] il peut faire les substitutions nécessaires, il ne peut plus faire les combinaisons. Vous voyez.
Ben, pour résumer très rapidement — parce que je ne voudrais là pas du tout m’étendre, il faut regagner le temps que j’ai perdu — je dirais ceci : qu’est-ce c’est le « trouble de contiguïté » d’après Jakobson — là je parle pour mon compte, je ne parle plus d’après Jakobson –. Jakobson distingue très, je reviens vite sur Jakobson, il distingue parfaitement les deux troubles [Pause] en disant que le « trouble de contiguïté », c’est-à-dire lorsque c’est le pouvoir de construction qui est troublé, [32 :00] c’est un trouble que porte avant tout sur le message. [Pause] Le « trouble de similarité », au contraire, c’est un trouble qui porte avant tout sur le code. [Pause] Ça, ça se comprend tout seul. Quand il est plus précis, parce qu’il y a une telle interrelation entre le code et le message, qu’il éprouve le besoin d’être en effet plus précis, et il nous dit que le trouble de similarité est d’avant tout un trouble de décodage, et que le trouble de contiguïté est avant tout un trouble d’encodage. Et en fait, tous ceux qui ont fait un tout petit peu de linguistique savent tout ça.
Moi, pour mon compte, j’en tire juste la conclusion suivante, c’est que [33 :00] le trouble de contiguïté, il est avant tout moteur — vous suivez, vous vous rapportez à mon schéma, « moteur », c’est « M », là, dans l’axe horizontal — il est avant tout moteur. En effet, il porte sur la construction des phrases prononcées, sur la combinaison des éléments. [Pause] Et il est avant tout aussi désintégrateur. Pourquoi ? C’est un trouble qui porte, on l’a vu, sur la combinaison, la combinaison devant [34 :00] s’entendre de deux manières [Pause] : combinaison d’une unité linguistique par de sous-unités, par des unités inférieures, [Pause] unités inférieures, par exemple, le phonème, les phonèmes comme sous-unités par rapport aux morphèmes, donc trouble de la combinaison d’une unité par des sous-unités, et de la combinaison par laquelle une unité entre dans des unités supérieures. C’est donc un trouble moteur et d’intégration. [Pause ; Deleuze se déplace vers le tableau] [35 :00] Ce qui m’arrange, ça me permettrait donc de tracer de « I » [Intégration] à « M » une ligne en pointillé, qui représenterait cette aphasie motrice désintégratrice. Et « aphasie motrice » est un terme courant dans la théorie de l’aphasie du 19ème siècle.
L’autre trouble, trouble de similarité, [Pause] lui, c’est un trouble essentiellement sensoriel [Pause] et de différenciation. [Pause] Pourquoi ? Précisément parce qu’il porte [36 :00] sur quoi ? Tout comme je disais tout à l’heure qu’il y a deux niveaux d’intégration — l’intégration qu’une unité opère par rapport à de sous-unités, et l’intégration dans laquelle elle entre par rapport à une unité supérieure — de même il y a deux aspects de la différenciation. Dans le langage, on le voit bien. Il y a un premier aspect de la différenciation qu’on pourrait appeler le discernement des traits distinctifs — je vais vite parce que ça n’a pas d’importance, là je parle uniquement pour ceux qui ont fait un peu de linguistique — un phonème est un porteur d’une pluralité de traits linguistiques, de traits distinctifs.
Si je donne quand même un exemple pour que ceux qui ne savent pas puissent suivre, par exemple je dis le mot « cochon », le mot « cochon », j’extrais le phonème [37 :00] « ch ». Eh ben, « ch » est, comme on dit, comme les linguistes disent, un faisceau de traits distinctifs. Pourquoi ? Vous avez « ch » sur « t », « cochon » sur « coton », vous avez « ch » sur « deux s », « cochon », « cosson », vous avez, cherchons, quoi ? [Pause ; Deleuze sollicite des suggestions de substitution] Colon, ah oui, « ch » sur « l », « cochon », « colon ». Chaque fois vous avez, il n’y a pas un phonème qui ne soit pas un ensemble de traits distinctifs. Donc le premier aspect de la différenciation, je dirais, pour mon compte, c’est la sélection des traits distinctifs, ou plutôt, non, le discernement, c’est le discernement des traits [38 :00] distinctifs. Et le deuxième aspect de la différenciation, c’est la substitution de deux formules qui diffèrent sous un aspect et se ressemblent… la substitution de l’une à l’autre de deux formules qui diffèrent sous un aspect et se ressemblent sous un autre. [Pause]
Dès lors, ça doit vous paraître évident que l’aphasie qui présente des phénomènes de dé-différenciation soit au même temps une aphasie sensorielle, tandis que l’aphasie qui présente des traits de désintégration est forcément une aphasie motrice. [Pause] Si bien que mes deux types d’aphasie, voyez, [Deleuze se tourne vers le tableau] je pourrais tracer [39 :00] une ligne allant de « I » à « M », c’est l’aphasie motrice de désintégration — là ce que je vous dis, ce n’est plus du Jakobson, ça ne fait rien, mais enfin ne lui faites pas dire ça, il ne serait pas content –, et l’autre aphasie « S » [Sensori] « D » [Différenciation], c’est l’aphasie sensorielle opérant par dé-différenciation. Alors là, ça serait très compliqué. Moi ce que je voudrais montrer, c’est que dans la théorie classique de l’aphasie, les autres types d’aphasie, moi ça me permettrait de remplir les autres choses, mais enfin ça ne nous donnerait que peu de satisfaction. Juste, donc, j’en reste là pour le moment. J’espère que vous retenez vaguement, vous retenez, que vous essayiez de retenir [40 :00] vaguement quelque chose.
Je dis, eh ben, en gros, c’est ça l’image classique du cerveau. Moi, je me suis servi, j’ai invoqué l’exemple de l’aphasie parce que, d’une part, ça me plaît de remarquer que des linguistes, vous savez ce qu’ils ont fait les linguistes ? Mais ils ont gardé absolument, dans cette discipline si nouvelle, ils ont gardé le plus vieux schéma cérébral. Alors je ne cite que les plus grands parmi les linguistes. Je viens d’essayer de le démontrer pour Jakobson, mais pour [Noam] Chomsky alors, il faudrait le montrer en détail. Ça nous ferait trois heures, mais ça ne fait rien. Chez Chomsky, c’est étonnant. D’ailleurs il le reconnaît lui-même ; quand il a fait son retour à Descartes, ce n’est pas par hasard. Le modèle cérébral, c’est ça qui m’intéresse, moi, chez les linguistes, c’est de savoir au juste quel est le modèle cérébral sous-jacent qu’ils [41 :00] emploient. Je crois que la linguistique a absolument besoin d’un modèle cérébral et qu’elle le suscite, qu’elle le suscite et qu’elle s’en sert implicitement. Alors quand on parlera un peu des problèmes au niveau du cinéma, par exemple, de la sémiocritique d’inspiration linguistique, une question principale, c’est, par exemple, quand on pense aux études très sérieuses, très importantes, très intéressantes de Christian Metz, c’est quel modèle cérébral implique sa conception du cinéma.
Et ce qui me paraît frappant en tout cas chez les linguistes, c’est à quel point ils sont accrochés à cette conception du cerveau : intégration-différenciation des centres, association sensorimotrice des éléments. Vous me direz, mais il n’y a pas de mal, mais je ne dis pas [42 :00] qu’il y ait du mal. Il n’y a aucun mal. Simplement, voilà une discipline nouvelle qui opère avec un vieux schème cérébral. Voilà. Je m’en réjouis uniquement par méchanceté pure. [Rires] Eh ben, ça me frappe beaucoup dans la conception de l’aphasie de Jakobson, elle est très, très forte. Vous lisez le livre, puisque tous ces cours, c’est pour vous donner envie de lire ou ceci ou cela. Vous verrez, vous verrez vous-mêmes, mais ce n’est pas par hasard qu’il retrouve exactement le schéma des aphasies, tel qu’il était proposé par les biologistes du cerveau du 19ème siècle. Voilà un premier point terminé.
Je dis, vous voyez, c’est ça, en gros… en gros, c’est ça, le cerveau comme arbre. [43 :00] Eh ben, on se vit comme ça, ou du moins, on s’est beaucoup vécu comme ça, ou bien nos pères se sont vécus comme ça. C’est marrant, imaginez de vivre… là, je parle du vécu, je saute au vécu. Je n’en dis pas grand-chose. Mais ce sont des gens qui se vivent avec un arbre dans la tête. Il y a beaucoup de gens. Et ce n’est pas par hasard que l’arbre est à la fois le modèle du cerveau et le modèle du savoir, de l’arbre de la connaissance à l’arbre du cérébral. [Pause] Bien, courte pause.
Qu’est-ce qui a changé ? Moi, je vois [44 :00] deux points de mutation, deux points de mutation, mais encore une fois, ça s’empiète tellement, c’est très délicat, très délicat. Je dirais aussi bien, oui, je précise, je reviens en arrière, aussi bien que système arborescent, je dirais de ce système cérébral que c’est un système centré. C’est un système centré puisque tout son axe vertical est fondé sur les centres et que l’ensemble de la circulation est fondé sur l’interaction des centres et des éléments. D’accord ? C’est donc un système arborescent et centré. [Pause]
Alors on passe à la recherche [45 :00] de notre mutation avec les précautions que j’ai prises. Faites attention, ça ne s’est pas fait en un coup ; ça s’est fait par-ci, par-là. Ben, reprenons l’axe intégration-différenciation, [Pause] et je me dis, et déjà dans la plus vieille biologie, et notamment avec le grand Claude Bernard, les problèmes d’intégration et de différenciation se sont pris, se sont trouvés inexplicablement liés avec un autre type de problème. Et ce n’est parce que les deux étaient très liés que c’était le même problème. L’autre problème qui venait recouper le thème de l’intégration et la différenciation, c’était [46 :00] le problème des milieux extérieurs et intérieurs. Plus le vivant était compliqué, complexe, plus il se constituait des milieux intérieurs. Plus ces milieux intérieurs étaient forts et structurés, plus ils dépendaient de mécanismes cérébraux [Pause] et de régulation cérébrale. Bien plus, la différenciation organique [Pause] et l’organisation des milieux intérieurs et extérieurs à chaque instant s’entrecroisent. Je veux dire, [47 :00] la distinction des milieux extérieurs et intérieurs favorise et accélère les processus de différenciation organique [Pause] et, inversement, les suppose. Là aussi, il y a une pleine interaction entre différenciation et constitution des milieux respectivement intérieurs et extérieurs.
Si bien que l’intégration et la différenciation se font comment ? Entre autres, elles se font par des niveaux relatifs d’extériorité et d’intériorité. Pourquoi je dis « relatifs » ? Parce qu’à première vue, il n’y a pas de milieux extérieurs [48 :00] et de milieux intérieurs absolus. Ce qui est un milieu intérieur d’un certain point de vue est un milieu extérieur d’un autre point de vue. Tout dépend de quelle unité organique vous prenez. Vous pouvez prendre évidemment l’organisme en son ensemble, mais même là, ça fera des difficultés. Prenez l’organisme en son ensemble, vous me direz, ce n’est pas compliqué : milieu intérieur, c’est ce qui est dedans. Comment se définit un organisme évolué ? Par ceci : c’est que précisément qu’il s’est constitué un milieu intérieur. Ça veut dire quoi il se constitue un milieu intérieur ? Ça veut dire se constituer en milieu qui ne dépend plus des variations des milieux extérieurs. Un milieu qui jouit d’une autorégulation. Par exemple, [Pause] la chaleur du sang [49 :00] en tant qu’elle devient indépendante des variations du milieu extérieur. Vous avez constitution d’un milieu intérieur.
Si vous prenez l’ensemble de l’organisme, vous pouvez dire eh ben, ce n’est pas compliqué ; le milieu intérieur, c’est tout ce qui est dedans. Pas sûr. Pas sûr. Qu’est-ce que vous diriez de l’estomac ? C’est un milieu intérieur ? Oui, c’est un milieu intérieur, jusqu’à un certain point. Par rapport à quoi il est intérieur ? Parce que par rapport à autre chose, peut-être qu’il est extérieur. Un milieu intérieur peut être d’un autre point de vue un milieu extérieur qu’on appellera un milieu extérieur annexé à l’organisme. À plus forte raison l’intestin. C’est un milieu intérieur, le milieu intestinal. Mais d’un autre point de vue, il joue le rôle de milieu extérieur annexé. [50 :00] Si bien que les déchets intestinaux seront franchement rejetés dans le milieu extérieur tout court.
Un texte de [Gilbert] Simondon insiste très bien sur ce point. Dans son livre, dans un livre très beau, L’individu et sa genèse physico-biologique [Paris : PUF, 1964 ; Grenoble : Millon, 1994]. Voilà ce qu’il nous dit, comme il le dit mieux ce que je veux dire, autant vous le lire : [Pause] « Il y a plusieurs étages d’intériorité et d’extériorité ; ainsi, une glande à sécrétion interne déverse dans le sang ou dans quelque autre liquide [51 :00] organique les produits de son activité : par rapport à cette glande » — par rapport à la glande à sécrétion interne – « le milieu intérieur de l’organisme général est en fait un milieu d’extériorité. De même, la cavité de l’intestin est un milieu extérieur pour les cellules assimilatrices qui assurent l’absorption sélective au long du tractus intestinal. … [Pause] L’espace des cavités digestives, c’est de l’extériorité par rapport au sang qui [52 :00] irrigue les parois intestinales ; mais le sang à son tour est un milieu d’extériorité par rapport aux glandes à sécrétion interne, qui déversent les produits de leur activité dans le sang » [Edition Millon, p. 224]
Donc, voilà tout ce que je veux dire. Et là aussi je n’ai pas le temps de développer, parce que … Voilà, l’hypothèse à laquelle je voudrais tourner, c’est que les processus d’intégration et de différenciation organiques sont strictement inséparables, des distinctions entre milieux intérieurs et milieux extérieurs, [53 :00] ces distinctions restant relatives, [Pause] étroitement relatives, ce qui veut dire qu’un milieu dit intérieur d’un certain point de vue sera extérieur d’un autre point de vue, qu’il y aura toute une chaîne des médiations intérieures et extérieures. Ces distinctions entre milieux intérieurs et milieux extérieurs restent relatives tant qu’on les rapporte à l’axe intégration-différenciation. Voilà, à la fois l’axe intégration-différenciation passe par ces distinctions, lesquelles sont relatives par rapport à lui. [Pause] D’accord, ça doit être très lumineux ça, hein, [54 :00] parce que…
Bon, alors. Ça n’empêche pas, ajoute Simondon dans des pages très, très étranges, mais que je trouve très belles, si on se rapporte à l’état de l’organisme le plus simple, l’organisme le plus simple… [Pause] — C’est ennuyeux tout ça, mais la conclusion va être si belle grâce à Simondon. — Si on se rapporte à l’organisme le plus simple, qu’est-ce qui se passe ? Lui, l’organisme le plus simple, c’est-à-dire, non différencié et non intégré, [Pause] il jouit [55 :00] d’une intériorité et d’une extériorité absolues, il jouit d’une intériorité et d’une extériorité absolues qui seront distinguées par quoi ? Par une membrane polarisée, l’expression la plus simple de la vie. Il y a l’extérieur et il y a l’intérieur. En d’autres termes, l’organisme le plus simple se dispose d’après un dehors et un dedans absolus. [Pause] À mesure qu’ils se compliquent, c’est-à-dire, s’intègrent et se différencient, les organismes passent par des niveaux relatifs d’intériorité et d’extériorité.
Reste la troisième conclusion, arrangez-vous comme vous pouvez : [56 :00] le cerveau restaure à sa manière un dehors et un dedans absolus [Pause] qui est comme la limite de toute relativité des milieux extérieurs et intérieurs. Qu’est-ce que ça veut dire ça, le cerveau, là ? Ben oui. Le cerveau va mettre en présence un dedans absolu et un dehors absolu. En présence, ça veut dire quoi ? En contact, en contact sans distance. C’est par-là que l’extérieur et l’intérieur ne sont plus relatifs. Le cerveau restitue un dehors et un dedans absolus, c’est-à-dire ce qui était la propriété de l’organisme le plus simple. [57 :00] Il le restitue à sa manière. Il met en contact un dehors absolu au-delà de tous les milieux extérieurs qui sera sans doute comme l’horizon de tous les milieux extérieurs, et un dedans absolu, au-delà de tous les milieux intérieurs, qui sera sans doute — comment dirai-je, pour parler romantique — le sans-fond de tous les milieux intérieurs, ou le plus profond.
Le cerveau va mettre en contact un dehors plus profond que tous les mondes extérieurs, un dedans – voyez où je veux en venir, si vous vous rappelez ce qu’on a fait précédemment – et un dedans plus profond que les mondes, que les milieux intérieurs. Comment qu’il fait cette mise en contact, une mise en contact sans distance du dehors et du dedans ? L’organisme élémentaire, lui, il avait besoin d’une distance entre son intériorité absolue et son extériorité absolue. Une mise en contact sans distance, ça veut dire quoi ? [58 :00] C’est ce que Simondon explique dans ces pages très belles. Ça veut dire, le cerveau a une structure topologique… [Interruption de l’enregistrement] [58 :10]
Partie 2
… Le cerveau a une structure topologique qui assure la co-présence d’un dehors et d’un dedans absolus. Vous me direz : qu’est-ce que c’est que ça ? Qu’est-ce que ça veut dire ? Eh ben oui, eh ben oui… Il faut signaler juste deux choses-là, à ma connaissance, même la biologie du cerveau ne rappelle pas, il a fallu que ce soit un philosophe très compétent en biologie, en technologie et en cybernétique, qui le rappelle un peu parce que j’ai l’impression que la cybernétique, elle s’en occupe beaucoup de ça. C’est que finalement, comme dit Simondon, le cerveau n’a pas d’interprétation euclidienne. [Pause] Le cerveau ne s’interprète pas en espace euclidien. [59 :00] L’intégration-différenciation s’interprète en espace euclidien, les milieux relatifs intérieur-extérieur, oui. Mais le cerveau, lui, il implique quoi ? Un espace topologique. [A ce propos, voir Simondon, L’individu et sa genèse physico-biologique, Édition Millon, p. 223, et aussi L’Image-Temps, p. 275, note 31]
Comme dit, là je cite un texte intéressant : « Le développement du néo-pallium dans les espèces supérieures se fait essentiellement par un plissement du cortex : c’est une solution topologique, non pas une solution euclidienne. … À la rigueur » — là je trouve ce texte très, très beau, peu importe, il ne faut pas comprendre trop, hein ? [Rires] — « À la rigueur. il ne faudrait pas parler de projection pour le cortex » — il fait allusion à ce dont je vous ai parlé tout à l’heure [60 :00] : les aires de projection sensorielles, c’est-à-dire, les aires de projection, les projections des organes des sens, dans des aires cérébrales – « À la rigueur, il ne faudrait pas parler de projection pour le cortex » — et, en effet, là il n’y a pas un biologiste qui ne sait que les projections supposées des organes des sens sur une aire cérébrale sont des schémas d’une grossièreté, d’une grossièreté insensée, quoi – « il ne faudrait pas parler de projection pour le cortex, bien qu’il y ait, au sens géométrique du terme, projection pour de petites régions ; il faudrait dire [plutôt] : conversion de l’espace euclidien en espace topologique. Les structures fonctionnelles de base du cerveau sont topologiques ; le schéma corporel convertit ces structures topologiques en structures euclidiennes à travers un système [61 :00] médiat ». Médiat, c’est-à-dire, à travers la médiation des milieux intérieurs et des milieux extérieurs relatifs. [Simondon, Édition Millon p. 225]
Et, qu’est-ce que ça veut dire, ça ? Qu’est-ce que ça veut dire ? Mettre en contact ou en co-présence un dedans plus profond que tous les milieux intérieurs relatifs, et un dehors plus profond que tous les milieux extérieurs relatifs. Ce serait ça. Ce serait ça la fonction topologique du cerveau. Voyez ? Ce qui lui permet de dire : « La structure non topologique d’intégration et de différenciation, la structure non topologique d’intégration et de différenciation apparaît comme un moyen de médiation et d’organisation » – de médiation et d’organisation [62 :00] grâce aux milieux intérieurs et aux milieux extérieurs relatifs, donc – « apparaît comme un moyen de médiation et d’organisation pour soutenir et étendre la première structure » — c’est-à-dire la structure topologique – « qui reste non seulement sous-jacente, mais fondamentale ». [Simondon, Édition Millon p. 224]
Alors, moi, là je ressens un très grand bonheur parce que c’est une confirmation, par de tout autres moyens, on n’a pas cessé de tourner — quand je ferai la récapitulation la prochaine fois, on verra — vous vous rappelez tout ce qu’on a vu sur cette mutation de la pensée, dans la mesure où elle devient une pensée du dehors, mais d’un dehors plus profond que le monde extérieur, et qui est aussi bien une pensée du dedans, mais un dedans plus profond que tout milieu intérieur. Et voilà qu’au niveau du cerveau, on retrouve exactement le même problème, au niveau d’une relation topologique entre un dehors et un dedans absolus. [63 :00] Concrètement, vous allez me dire, que vous suppliez un petit concret, ça veut dire quoi ? Ça veut dire une chose très simple. Ça veut dire : plus un organisme est évolué et soumis à un cerveau structuré, [Pause] plus la totalité de son passé, [Pause] c’est-à-dire le dedans absolu — le dedans absolu, c’est précisément le Tout du passé –, [Pause] plus la totalité de son passé est mise en contact topologique, sans distance et sans retard — la topologie ignore les distances –, sans distance et sans retard [Pause] [64 :00] avec l’ouverture du futur, c’est-à-dire, avec le dehors absolu. [Pause]
Comprenez ce que ça veut dire. Mais c’est bien comme ça dans le cas du vivant. C’est même ce qui distingue le vivant et le cristal. Quand un cristal croît, qu’est-ce qui compte ? C’est les couches moléculaires : elles sont relativement intérieures et extérieures. C’est-à-dire, sera dite intérieure la couche moléculaire déjà constituée, sera dite extérieure la couche moléculaire en voie de constitution. Eh ben, vous pouvez vider le cristal de la majeure partie de sa substance intérieure, ça ne change rien, ça ne l’empêche pas de croître. Au contraire, un organisme, pas question que vous le [65 :00] vidiez de sa substance intérieure. Sa substance intérieure concentre, condense tout son passé. Et c’est tout son passé, c’est le Tout de ce passé, qui est en contact immédiat, c’est-à-dire topologique, qui est — comment dirait-on en topologie ? — qui est en voisinage, indépendamment de toute distance, qui est en voisinage avec le dehors absolu, c’est-à-dire, avec l’horizon du monde extérieur.
Si bien qu’il faudrait dire du cerveau qu’il reconstitue topologiquement [Pause] les conditions ordinaires de l’organisme le plus simple. Mais justement, pour les retrouver topologiquement, il faut la plus haute complexité. [Pause] [66 :00] Coprésence que — quitte à justifier plus tard, on a toute l’année pour ça, hein ? — coprésence que je pourrais dire, je pourrais dire dès lors que le cerveau, en tant qu’interprété topologiquement — et il doit s’interpréter topologiquement, et il doit s’interpréter topologiquement — le cerveau se présenterait comme le « contact sans distance » ou la « coprésence » : du dehors et du dedans, du vide et du plein, du passé et du futur, de l’envers et de l’endroit, etc. [Pause]
Il faut garder ces formules, là, du vide et du plein. Vous sentez que là j’ai déjà une arrière-pensée [67 :00] cinématographique. Qu’est-ce qu’il y a de plus plein qu’une image vide au cinéma ? Un espace vide de [Michelangelo] Antonioni, c’est une image pleine, une image de ciel vide de [Jean-Luc] Godard est une image pleine. Pleine de quoi ? [Interruption de l’enregistrement] [1 :07 :19]
… défini plus selon un axe, on ne le définira plus… je ne veux pas dire que ça s’exclut, il n’y a pas l’un qui est faux et l’autre… Mais ça n’est plus un axe d’intégration-différenciation qui sera fondamental, ce sera une « coprésence » d’un dehors absolu et d’un dedans absolu, d’un vide et d’un plein, d’un envers et d’un endroit, etc. [Pause] [68 :00] Si vous le vivez comme ça — même, il n’y a pas besoin de comprendre — si vous vivez votre cerveau comme ça, si vous le vivez topologiquement, et non plus sous les espèces de l’intégration et la différenciation, ben sentez que vous n’avez pas du tout le même rapport avec lui, oui ? Est-ce que vous pouvez le vivre comme ça ? Hein, eh ben, il faut voir, hein ? Mais ce n’est pas non plus facile. Est-ce que vous tenez à vivre sous forme intégration-différenciation ? Bon. Est-ce que c’est les mêmes trous qu’il y aura dans les deux cas ? Quelle répercussion pour une conception de l’aphasie ? N’allons pas trop vite.
Je dis, vous sentez, ça ne suffit pas. Il faut une seconde, une seconde petite mutation. Une seconde petite [69 :00] mutation parce que, sinon… Heureusement elle va être bien plus simple, parce que la première, elle était compliquée. Pour ceux qui veulent voir dans cette direction, mais je trouve que…, voyez Simondon, L’individu et sa genèse physico-biologique, page… le groupe de pages, c’est à partir de 257 suivantes, hein ? [Dans l’Édition Millon, la section « Topologie et ontogenèse » correspond aux pages 222-227] Je dis, je viens de m’attaquer, vous voyez, [Deleuze indique de nouveau le schéma au tableau] à mon axe vertical en disant : le vieil axe vertical intégration-différenciation, on va le remplacer par un axe topologique. Vous me suivez ? [Pause] Je dis : il me faut une seconde mutation parce que [70 :00] il faut s’attaquer maintenant à l’axe « S »-« M », sensorimoteur, à l’axe associatif. [Pause] Donc, ça se fait tout seul.
L’axe sensorimoteur, il se présente comment ? Il se présente sous forme de la cellule cérébrale, ou nerveuse, qu’on appelle « neurone ». Un neurone — je vous le donne, pour ceux qui n’ont…, je vous rappelle le vocabulaire de base, hein ? Je le prends au niveau vraiment le plus vulgaire des termes — le neurone, c’est la cellule, c’est la cellule cérébrale ou nerveuse. Cette cellule a des prolongements ramifiés extraordinaires. Ces prolongements ramifiés [71 :00] extraordinaires, on les appelle des « dendrites », D-E-N-D-R-I-T-E. Vous avez votre cellule nerveuse, votre neurone, et puis il a des prolongements qu’on appelle des dendrites. Les dendrites, ce sont donc des prolongements ramifiés qui reçoivent l’influx, qui reçoivent l’influx nerveux. [Pause] Si vous voulez avoir tout de suite une idée de la complication de la chose, c’est que c’est fou ce qu’un neurone a de dendrites, hein ? Il en a beaucoup, beaucoup. Mais si ce n’était que ça. Il y en a qui vont très loin, [72 :00] qui vont extrêmement loin. [Pause] Bon, ça se complique.
Et puis, enfin, vous avez, troisième grand élément, l’ « axone », l’axone, l’axone : A-X-O-N-E. L’axone, c’est aussi un prolongement. Mais cette fois, c’est un prolongement unique du neurone, prolongement unique qui se ramifie à la fin — alors, il peut se ramifier beaucoup, hein ? — qui se ramifie à la fin et qui, lui, transmet l’influx. Transmet l’influx à quoi ? On peut y répondre : aux dendrites du neurone suivant, du neurone suivant. Ça dépend. Ou pas du neurone suivant, ce n’est pas collé puisqu’il [73 :00] y a des dendrites qui vont chercher très loin. Rendez-vous compte donc déjà que ça ne va pas tout seul, qu’un de vos neurones mais est soumis à des influx parfois très lointains ? Il y a une multitude d’influx là en même temps ; c’est, c’est un système. Voilà.
Pendant longtemps l’image classique du cerveau a dit que tout cela formait un réseau continu. [Pause] Mais assez vite on s’est aperçu que ce réseau n’était pas continu, au sens classique du mot, mais était plein de coupures. [Pause] [74 :00] Ces coupures s’établissaient, par exemple, notamment entre la terminaison de l’axone du neurone « A » et la terminaison de la dendrite correspondant du neurone « B ». Vous vous rappelez ? Dendrite saisie reçoit l’influx, le transmet dans la cellule, dans le neurone, laquelle l’envoie dans l’axone qui va le repasser aux dendrites de la cellule suivante. Or, entre la terminaison de l’axone de la [75 :00] cellule « A » et la dendrite de la cellule « B », il y a coupure. Le réseau n’est pas continu.
Mais voilà que pendant longtemps, la coupure a été conçue, si l’on peut dire, de manière électrique, c’est-à-dire, d’un bord à l’autre de la coupure, la transmission de l’influx était électrique. [Pause] Cette coupure, elle reçoit un nom, c’est ce qu’on appelle une « synapse ». [Pause] [76 :00] C’est une synapse, c’est-à-dire c’est la communication, dans le cas le plus simple, de la fin d’un axone et du début de la dendrite de la cellule suivante. C’est donc, à la lettre, je peux dire du synapse, que c’est un point-coupure. C’est pour ça que très souvent dans les manuels, vous verrez la synapse définie comme un point de jonction : c’est le point de jonction de deux neurones. C’est un point ou une coupure. Ce n’est pas fait pour nous étonner, vu ce qu’on a vu, vu ce qu’on a fait précédemment. Nous disons, nous, c’est un point-coupure. [Pause]
Alors, tant qu’il y a eu la transmission, l’hypothèse de la transmission électrique, [77 :00] tout allait bien. Tout allait bien ou relativement bien. Il n’y avait déjà plus de réseau continu, mais enfin l’influx pouvait facilement franchir le point-coupure. Pourquoi ? Parce que, au moment du passage de l’influx, l’hypothèse était la suivante : les deux membranes cellulaires, c’est-à-dire, membrane de la dendrite et membrane de l’axone, plutôt l’inverse : membrane de l’axone et membrane de la dendrite suivante, se rapprochaient, devenaient adjacentes, devenaient adjacentes, ou bien même, suivant une hypothèse particulièrement audacieuse, ou bien même, formaient une seule et même membrane. Que ce soit adjacent ou une seule et même membrane, là, à nouveau nous avons [78 :00], nous avons toutes raisons de nous réjouir car nous sommes en droit, en vertu de ce qu’on a fait précédemment, de dire, sans aucunement forcer les choses, à mon avis : dans un tel cas, c’est un point-coupure rationnel.
Je dis : sans forcer les choses, c’est-à-dire sans aucune métaphore mathématique facile — puisque si vous vous rappelez, ça c’est la base, il faut vous rappeler, sinon c’est la catastrophe ; ce que nous appelions sur une droite, sur une droite continue, ce que nous appelions un « point-coupure rationnel », c’était un point qui opérait une répartition de deux ensembles, [Pause] mais tel que ce point [Pause] [79 :00] était ou bien la fin du premier ensemble ou bien le début du deuxième ensemble, c’est-à-dire, appartenait à l’un ou l’autre des deux ensembles, ou appartenait aux deux à la fois. Si on me dit : dans une transmission électrique, les deux membranes deviennent adjacentes, je dis : il y en a une qui marque la fin du premier ensemble et l’autre qui marque le début du deuxième ensemble. C’est typiquement un point-coupure. La synapse est un point-coupure rationnel. Si on me dit : une seule et même membrane se forme, je dis très bien, le point-coupure appartient aux deux ensembles : c’est un point-coupure rationnel. Voilà !
Ça me donne le troisième caractère [80 :00] de l’image classique du cerveau [Pause] : processus d’intégration-différenciation s’exerçant par rapport, deuxième caractère, s’exerçant par rapport à des enchaînements sensorimoteurs, les points, troisième caractère, les point-coupures de ces enchaînements étant des points rationnels, c’est-à-dire, les synapses étant des points rationnels. J’en ris tout seul parce que je vous soumets à un régime à la fois très désagréable de passer des mathématiques rudimentaires à de la biologie rudimentaire, [81 :00] mais je voudrais que vous sentiez que tout ça c’est uniquement pour faire des concepts en philosophie quoi, c’est du matériau… Vous me suivez ? — Je dirais, voilà les trois caractères, celui qui me manquait, vous voyez ? — Il faut…, c’est curieux comme tout ça est source de contentement profond. — On a un troisième caractère, qui nous manquait, ben on le tient maintenant. On a réintroduit dans le cerveau, ou dans l’arbre du cerveau, les point-coupures rationnels. Et voilà, et ça nous fait une image d’ensemble du cerveau comme arbre.
Si bien que ce fut une grande mutation peut-être, lorsque des doutes naquirent [82 :00] sur, non pas l’existence ou non d’un réseau cérébral continu — ça, la question est réglée, les synapses étaient prouvées, personne ne pouvait discuter l’existence des synapses — mais sur le mode de fonctionnement des synapses. Et lorsque les doutes se sont multipliés sur l’hypothèse de la transmission électrique, [Pause] et lorsqu’une toute autre hypothèse a surgi : l’idée d’une transmission chimique. [Pause] Ça veut dire quoi ? Ça veut dire que [83 :00] — vous allez tout de suite comprendre la différence — lorsque l’influx arrive à l’extrémité du neurone de la cellule « A », du neurone « A », eh ben, il n’y a ni adjacence de membranes, ni unité de membranes. Il se trouve devant une fente, il se trouve devant une fente à franchir. Imaginez, vous dites : ça commence à s’animer là, que notre cerveau va se peupler, de quoi ? Il va se peupler de millions de millions de millions de petites fentes, de fentes synaptiques. Bon, vous me direz : il était déjà, il y avait déjà des coupures. Bon, d’accord. [84 :00] Qu’est-ce qui se passe ? Comme dit un très bon auteur : un vide critique. Entre deux neurones, un vide critique.
Comment l’influx va se… Si l’influx n’a pas le pouvoir de se transmettre électriquement, comment il va faire ? Se transmettre électriquement, en science, ça veut dire, n’est-ce pas, avec des charges d’ions. Si ce n’est pas ça le mécanisme de la synapse, comment il va faire ? Ben, voilà qu’on se met à supposer que — on a des raisons pour ça — voilà qu’on se met à supposer que l’influx, quand il arrive à l’extrémité du neurone, va libérer quelque chose qui n’appartient ni à lui, l’axone de [85 :00] « A », ni à l’autre, c’est-à-dire à la dendrite de « B ». Il va libérer une substance chimique plus ou moins voisine de l’hormone, des hormones : un médiateur chimique, [Pause] un médiateur chimique. Encore faudrait-il savoir d’où il vient, ce médiateur chimique. [Sur cette discussion, voir L’Image-Temps, pp. 274-277, et surtout la note 32, p. 275]
Heureusement, avec les progrès de la microbiologie, on avait repéré dans la fente synaptique, dans la coupure synaptique, on avait repéré de bizarres vésicules, de petites vésicules. Alors, vous voyez ? De là [86 :00] à supposer que ces vésicules contiennent le médiateur chimique et que quand l’influx arrive au bout de l’axone du neurone « A », quand l’influx arrive au bout de son chemin, la vésicule va libérer une quantité du médiateur chimique. Ça devient de plus en plus intéressant, parce que si c’est des vésicules qui libèrent, la libération du médiateur ne se fera pas de manière continue non plus. On va avoir des discontinuités partout. Une vésicule contiendra une quantité très petite de médiateurs, de médiateurs chimiques de type hormonal. Elle va le lâcher. [87 :00] Donc le médiateur chimique sera lâché par quantités discontinues correspondant aux différentes vésicules qui le libèrent. C’est pour ça que les biologistes du cerveau vont parler de « quanta ». C’est par quanta discontinus que le médiateur chimique opérera. Et c’est par l’intermédiaire du médiateur chimique que l’influx passera du neurone « A » au neurone « B ».
Qu’est-ce que je vais en tirer ? Une chose toute simple, je n’ai pas besoin de… Le point-coupure, la synapse, reste un point-coupure. [Pause] [88 :00] Ce point-coupure détermine deux ensembles : le neurone « A », avec son axone et ses dendrites, le neurone « B », avec ses dendrites et son axone. Le point-coupure détermine deux ensembles. Seulement voilà, il ne fait plus partie ni de l’un, ni de l’autre des ensembles qu’il détermine : il est autonome. Ce n’est pas par métaphore ; c’est littéralement qu’on appelle cela une « coupure irrationnelle », c’est-à-dire une coupure qui détermine deux ensembles distincts, et qui pourtant ne fait partie ni de l’un, ni de l’autre des deux ensembles. [89 :00] Je dirais que dans le cas de synapses chimiques… alors que dans le cas des synapses électriques, nous nous trouvions devant des points-coupures rationnels, dans le cas des synapses chimiques, nous nous trouvons devant des points-coupures irrationnels.
Bon. Et alors, et alors, c’est tout le régime du cerveau qui passe, quoi, sous un régime, à la lettre, on va voir à quoi ça nous entraîne, sous un régime probabilitaire. Sous un régime probabilitaire, c’est-à-dire, quelles sont les probabilités pour qu’un message [90 :00] ou un influx arrivant par l’axone de « A » franchisse le vide critique, une fois dit qu’en même temps, toutes sortes d’autres messages, toutes sortes d’autres influx parviennent par d’autres dendrites ? Plus la synapse est chimique, plus le cerveau cesse d’être un système déterminé pour devenir un système probabiliste. Un Américain a intitulé un article — comme d’habitude, vous pardonnez mon anglais — « an uncertain system » : un système [91 :00] d’incertitude, un système probabilitaire. Mettons, pour le moment… on va voir que, justement, ce n’est pas ça, mais on progresse doucement. Un système probabilitaire, bien. [Deleuze cite l’expression « uncertain system » dans L’Image-temps, p. 275, note 32, sans y fournir de source sauf Jean-Pierre Changeux, L’homme neuronal (Paris : Fayard, 1983)]
Dès le moment où il y a un vide critique à franchir, vous êtes forcé de passer à une espèce de mécanisme aléatoire qui est le mécanisme du médiateur chimique et du nombre de quanta qu’il va lâcher. Une fois dit que le cerveau est assiégé de messages qui viennent de toutes ces dendrites, qu’il est perpétuellement, qu’à chaque fois, il y a coupure sous forme de points-coupures irrationnels, vous tombez dans la question [92 :00] : comment se font les enchaînements ? Comment se font les enchaînements d’un neurone à l’autre ? La réponse, c’est : les enchaînements ne sont plus déterminés, comme ils l’étaient sur le mode électrique. Les enchaînements deviennent probabilitaires. Qu’est-ce qu’un enchaînement probabilitaire ? Qu’est-ce que ça veut dire ça, un enchaînement probabilitaire ?
Pour le moment, retenons juste deux conclusions : les deux points de mutations dans le modèle cérébral se fait sous quelle forme ? Le cerveau implique, ou le cerveau promeut — oui, c’est encore mieux — promeut un espace, ou non, développe, oui, développe — non… enfin, peu importe, n’importe quel mot [Rires] — développe un espace topologique [Pause] [93 :00] et enveloppe un espace probabilitaire. Donc à l’axe vertical intégration-différenciation, nous substituons le contact topologique du dehors et du dedans. À l’axe horizontal sensorimoteur, nous substituons le caractère probabilitaire des enchaînements, puisque les enchaînements sont interrompus par des points-coupures irrationnels. De ces deux manières, nous ne pouvons plus [94 :00] — alors, pensons à la vie — nous ne pouvons plus vivre notre cerveau comme un arbre, nous n’avons plus un arbre dans la tête : c’est fini. C’est fini.
Un spécialiste du cerveau…, vous trouverez beaucoup de renseignements, par exemple, dans le livre connu de Jean-Pierre Changeux : L’homme neuronal [1983], sur les synapses, et vous trouverez aussi des renseignements sur, alors insistant davantage sur l’aspect probabilitaire du cerveau, chez un neurobiologiste américain [au fait, anglais] qui s’appelle Steven Rose, « Rose » comme une rose, dont le livre Le cerveau conscient [The Conscious Brain (New York : Knopf, 1973)] a été traduit en français il y a quelques années aux éditions du [95 :00] Seuil. Et dans le livre de Steven Rose, il y a là une métaphore qui est très intéressante. Il dit : vous comprenez, il y a des cas très spéciaux où un axone — parce qu’il essaye de dire dans chaque cas quelles sont les chances de probabilité pour qu’il y ait passage du message ou de l’influx — il dit : ça varie d’après plusieurs facteurs. Il y a, par exemple, des cas où le dendrite du neurone « B »… la dendrite, pardon, du neurone « B » s’enroule, s’enroule autour de l’axone. Vous voyez ? Ça s’enroule. Or les dendrites ont constamment des épines. Si bien qu’à chaque épine de la dendrite, il y a [96 :00] une jonction synaptique, il y a synapse. [Deleuze se corrige ici sur le genre du mot « dendrite » qu’il utilise dans cette séance tantôt au masculin, tantôt au genre correct, au féminin]
Donc, pour une seule dendrite et un axone, vous pouvez avoir une multiplicité de synapses dans ce cas, lorsque la dendrite avec ses épines s’enroule. Et la métaphore, c’est alors — aujourd’hui c’est un peu morne, tout ce que je dis, mais c’est en même temps source de petites joies constantes — il dit quelque chose de merveilleux, Steven Rose. Il dit : « c’est exactement comme lorsque » — attendez que je n’oublie pas la citation – « comme lorsque le liseron, le liseron s’enroule autour de la ronce ». C’est beau ça ! Et ça va nous servir, parce que ce n’est pas une métaphore. Le liseron, vous voyez, c’est de la mauvaise herbe. Le liseron [97 :00] s’enroule autour de la ronce, la ronce, c’est exactement la dendrite avec ses épines. Si je vous faisais un dessin des dendrites, mais enfin vous pouvez l’imaginer, un prolongement avec des épines. Voilà que cette dendrite particulièrement bizarre s’enroule autour de l’axone du neurone précédent ; à chaque contact d’épine, il y a une synapse. Donc, la dendrite, c’est la ronce, et l’axone s’enroule, est comme le liseron qui s’enroule autour de la ronce. Mais comme nous sommes loin du modèle de l’arbre ! On est dans un tout autre domaine.
En d’autres termes, sentez, sentez le monde futur, ce n’est pas un arbre que nous avons dans la tête, ce que nous avons dans la tête, [98 :00] c’est de l’herbe ! [Rires] Et il faut vivre comme ça. Et c’est par rapport vécu avec le cerveau. Je dis, par « rapport vécu », je pense à un grand auteur : Henry Miller. Il a un don prodigieux : il a toujours vécu son cerveau comme de l’herbe. Très curieux, mais ce n’est pas une image comme ça. Le dedans de sa tête, il le vit comme rempli d’herbe, quoi. Les Américains, les Américains, ils sont très doués pour ça. Les Américains, c’est très connu, ils ne croient pas aux arbres, ils croient à la prairie, alors… [Rires] Bon, ils ont toujours vécu ce qui se passait dans leurs têtes comme de l’herbe, depuis l’un des plus grand poète, depuis le, depuis le poète fondateur [99 :00] de l’Amérique, ça a été comme ça. [Sans doute, Walt Whitman, Leaves of Grass]
Mais c’est très curieux. Et lorsque, et lorsque Miller se dit : « moi, sous quelle forme est-ce que je renaîtrai ? », parce qu’il fait semblant de croire ou il faisait semblant de croire aux renaissances, il disait « sous forme d’un parc, je renaîtrai sous forme d’un parc ». Il y a des gens qui veulent renaître sous forme de brin d’herbe, et puis il y a de gens qui veulent renaître sous forme d’arbre, c’est leur affaire. Mais à ce moment-là, il ne faut pas qu’ils aient les mêmes synapses, hein ? Parce que si on marche sur des synapses chimiques, l’arbre-cerveau, c’est fini. On peut espérer tout au plus d’assister, même de favoriser, les noces d’une ronce et d’un liseron. Et quoi de plus beau ? Alors, ou bien vous préférez la force d’un peuplier et vous renaîtrez peuplier, ce que vous êtes déjà [100 :00] ; ou bien vous aimerez les noces du liseron et de la ronce. À votre choix ! Mais dans les deux cas, vous ne penserez pas pareil, hein ? Mais il y aura quand même des possibilités de se comprendre ! Voilà ! Bon.
Alors, je tire là mes conclusions provisoires. C’est bien, encore une fois, un tout autre schéma qu’on se trouve devant un modèle cérébral, où encore une fois, l’axe horizontal, [Pause] l’axe horizontal est devenu probabilitaire et l’axe vertical est devenu topologique. Dès lors, il ne s’agit plus d’un mécanisme d’intégration-différenciation. Il s’agit du dehors et du dedans. Passant par la distribution relative du milieu intérieur et du milieu extérieur. Et dans l’autre cas, il ne s’agit plus d’enchaînement [101 :00] sensorimoteur ; il s’agit de quoi ? Il s’agit de communication probabilitaire, et on se trouve devant un problème — avant de se reposer un petit peu – eh ben, comment peuvent se faire, et comment concevoir des telles communications probabilitaires ? Comment peuvent se faire… qu’est-ce que c’est ce mode d’enchaînement qui n’est plus un enchaînement de type sensorimoteur ? Voilà où nous en sommes. On se repose un peu ? Pas longtemps, vous comptez six minutes, hein ? Six minutes … [Interruption de l’enregistrement] [1 :41 :43]
… la formule, à la recherche d’une formule concernant un enchaînement qui ne serait plus du type sensorimoteur. [102 :00] Alors, vous voyez que, quand même, ça se tient avec tout ce qui précède puisque tout ce qui précède était fondé sur une rupture, et sur l’existence d’une rupture sensorimotrice. Donc, en cas de rupture sensorimotrice, comment concevoir les enchaînements, et sous quelle forme ? À moins qu’il n’y ait plus du tout d’enchaînements, mais bon. Eh ben, là, je vais procéder comme… Je voudrais multiplier les directions de recherche et vous proposer quatre ou cinq directions de recherche très différentes, donc.
Première direction : [Pause] il y a des choses bizarres qui se passent, je veux dire, dans des domaines, appelons-les pour le moment, des « domaines intermédiaires ». [103 :00] Intermédiaires entre quoi et quoi ? Entre le hasard et la détermination, le hasard et la dépendance. Qu’est-ce que c’est que ça ? Exemple de hasard : une loterie [Pause] avec des tirages au hasard successifs, ces tirages étant indépendants les uns des autres. La condition même est que les tirages soient indépendants. De même, le jeu de la roulette : les lancés sont indépendants les uns des autres. [104 :00] On peut toujours dégager, d’un tel domaine du hasard, on peut dégager des lois dites de probabilité, bien. Je mélange tout : ces lois dites de probabilité sont bien connues et soumises à certaines conditions : l’indépendance des particules ou des tirages, [Pause] la fluidité de l’espace ambiant, la vitesse de diffusion, etc., [Pause] et son lien-domaine qu’on connaît bien dans la théorie cinétique [105 :00] des gaz, avec le thème du mouvement brownien, mouvement brownien de particules dans le fluide. Voilà. Puis il y a, d’autre part, des enchaînements déterminés. [Pause]
Bien, qu’est-ce qu’il peut y avoir entre les deux ? Tout un domaine a été identifié, encore fallait-il reconnaître ce domaine, sous le nom d’un domaine de « phénomènes aléatoires partiellement dépendants », ou phénomènes semi-aléatoires, phénomènes aléatoires partiellement dépendants, [106 :00] donc qui ne se ramènent ni sur le phénomène semi-aléatoire… des « mixtes » de dépendance et d’aléatoire. Première direction : comment obtenir un tel mixte d’aléatoire et de dépendant ? Opérer par tirages au hasard, mais tirages au hasard successifs qui dépendent les uns des autres. Qu’est-ce qui se passe ? C’est un domaine mathématique, ça. Qu’est-ce qui se passe dans le cas de tirages au hasard successifs mais dépendants les uns des autres ? C’est ce qu’on appellera des « phénomènes semi-aléatoires », des phénomènes aléatoires partiellement [107 :00] dépendants… [Interruption de l’enregistrement] [1 :47 :02]
Partie 3
… Ben, dans une loterie, vous faites un tirage, un premier tirage, A, un second tirage, B, un troisième tirage, C. Chacun de ces tirages est strictement indépendant des autres. Au point que, au bout d’un certain temps, vous devez changer celui qui tire pour qu’il n’y ait même pas de dépendance musculaire de la part du tireur. Imaginez l’autre situation. Une drôle de chaîne, ça va faire une chaîne. Une chaîne de tirage au hasard, mais chaque tirage au hasard dépend de celui qui précède. [Pause] Une telle chaîne, [108 :00] elle est devenue bien connue en mathématiques, en cybernétique, sous le nom de l’un de ses créateurs : chaîne de [Andrei] Markov — M-A-R-K-O deux F ou V à votre choix — chaîne de Markov. Et pour mieux comprendre, là, je donne un exemple rapide, mais voilà.
Markov avait commencé par étudier une drôle de chose : comment on peut faire du faux latin ou du faux anglais, du faux français ? Comment on peut fabriquer une fausse langue ? [Pause] C’est bien connu sous le nom de « problème des jargons ». Comment faire du jargon ? [Pause] [109 :00] Vous pouvez toujours concevoir… alors vous vous donnez une série d’urnes là, des urnes, des boîtes. Je suppose que je me donne la règle suivante : chaque boîte va contenir des groupes de trois lettres, [Pause] dont les deux premières sont constantes, [Pause ; Deleuze chuchote à quelqu’un près de lui] dont les deux premières sont constantes. Alors dans votre première urne, vous mettez a-b-a, a-b-b, a-b-c, a-b-d, a-b-e, a-b-f, etc. Vous comprenez ? Dans la seconde, vous mettez [110 :00] b-c-a, b-c-b, b-c-c, b-c-d, etc. Donc chaque urne contient un groupe de, on appellera ça, de « trigrammes », groupe de trois lettres dont les deux premières sont constantes. Vous me suivez ?
Si vous faites ça, ça ne donne rien encore. Mais vous savez bien que sur une langue, il est capable de faire des calculs de fréquence, par exemple, au niveau des groupes de lettres. On dira en français que la lettre Q, dans un nombre maximum de cas, est suivie par U, [111 :00] et on pourra calculer la fréquence du groupe Q-U par rapport à des groupes comprenant Q qui ne serait pas suivi de U. Là immédiatement, je ne vois que COQ, bon. De même en français, il y a une fréquence importante où la lettre H est précédée par C. Vous pouvez calculer — et là les ordinateurs servent beaucoup, évidemment, c’est pour ça que tout ça était très lié à des problèmes d’ordinateurs, et que les chaînes de Markov ne pouvaient guère être découvertes avant — vous pouvez établir la fréquence de C-H. Donc supposons que vous fassiez des calculs de fréquence sur une langue, ça a été très souvent fait. Vous pouvez faire aussi des calculs de fréquence [112 :00] sur la langue d’un auteur. Ça a été fait notamment pour [James] Joyce, dans des conditions très intéressantes. Vous pouvez faire des calculs de fréquence pas seulement sur des groupes de lettres, mais sur des mots, ça a été fait pour [Stéphane] Mallarmé, bien. On ne sait pas bien où ça mène tout ça… Si, ça va permettre de faire des jargons, des faux Mallarmé, des faux Joyce.
Mais comment vous allez faire ? Vous prenez un seuil de fréquence qui vous assure donc que votre trigramme, c’est-à-dire votre groupe de trois lettres, a une certaine fréquence dans la langue considérée. Voilà. Et puis vous y allez. [Pause] Vous tirez d’une urne, vous prenez une urne, vous prenez l’urne, chaque urne donc est définie par [113 :00] les deux premières lettres des trigrammes. Vous prenez la lettre, vous commencez par la lettre, par l’urne I-B ; dans l’exemple là que j’ai sous les yeux, tiré d’un livre de [Raymond] Ruyer dont je vous parlerai… [Interruption de l’enregistrement] [1 :53 :22]
… et là, dans le cas qui nous occupe, on tire U. On écrit donc IBU. Le premier tirage vous a donné IBU. C’est un tirage de loterie. IBU, I-B-U. Là-dessus, vous avez deux cas : ou bien vous allez prendre n’importe quelle urne et faire un nouveau tirage, à ce moment-là, ce sera [114 :00] une succession de tirages au hasard indépendants ; ou bien — vous avez déjà deviné, j’espère — vous allez prendre l’urne B-U, et vous tirez. Supposons que vous sortiez BUS. De l’urne I-B, vous avez sortis IBU ; de l’urne BU, B-U, les deux dernières lettres, vous avez sorti BUS. [Pause] Il vous reste à prendre la boîte US, U-S, [115 :00] dont vous allez sortir U-S-C, hein ? U-S-C. Puis de la boîte S-C, vous allez sortir S-C-E. [Pause] Bon, vous avez une suite ; là-dessus évidemment vous vous donnez le droit de la couper. Ce qui vous donne, je vous lis lentement — je fais appel particulièrement à ceux qui ont fait un petit peu de latin — IBUS, I-B-U-S, coupure, vous coupez là ; CENT, C-E-N-T, [116 :00] coupure ; vous allez continuer vos tirages, IPITIA, I-P-I-T-I-A, coupure ; VETIS, V-E-T-I-S, coupure, etc.
Comme on dit, en gros, ça a l’air incontestablement d’être du latin, et vous voyez pourquoi, ce n’est pas compliqué, ce serait étonnant, puisque vous avez retenu des trigrammes d’après des fréquences, d’après des fréquences statistiques effectuées sur l’ensemble de la langue. Vous pouvez faire de même du faux anglais ; c’est ce qu’on appellera un jargon. Vous avez fait du jargon latin. Vous pouvez de la même manière faire du jargon [117 :00] joycien. Même ce jargon latin là, dans cet exemple, l’exemple est particulièrement développé par, dans un livre justement d’un linguiste qui s’appelle [Pierre] Guiraud, Les caractères statistiques du vocabulaire [Paris : PUF, 1954] Bon, vous pouvez fabriquer des jargons. Qu’est-ce qu’il y a ? Pourquoi je raconte tout ça ? Je vous raconte parce que vous avez le cas typique ; ça doit vous faire comprendre ce qu’on appelle une « chaîne de Markov », plus que si je la définissais par des formules mathématiques, vous voyez que chaque tirage se fait au hasard, dans l’urne, mais chaque tirage en même temps dépend du tirage précédent, puisque vous avez choisi là comme seconde urne, celle qui était déterminée par les deux derniers mots [lettres] [118 :00] du trigramme choisi dans la première urne.
C’est exactement ce qu’on appellera des phénomènes aléatoires semi-dépendants, [Pause] ou des phénomènes aléatoires partiellement dépendants. Il y a chaque fois tirage au hasard, mais dans des conditions de dépendance entre les urnes et les tirages successifs. Vous n’êtes donc pas dans les conditions de la loterie ; vous n’êtes pas non plus dans les conditions d’un déterminisme. C’est du semi-aléatoire, c’est une chaîne Markov. Vous pouvez dire qu’à la lettre, c’est un mixte, c’est un mélange, quoi, c’est un mélange structuré. [119 :00] On dira que c’est un mélange structuré en ce sens qu’il n’est pas structuré comme la langue et que pourtant ce n’est pas un mélange au hasard. [Pause]
Deuxième direction de recherche, pour aller vite. À première vue, ça n’a rien à voir. Dans la cosmobiologie, dans l’éternel problème de comment, comment est-ce que un organisme vivant a-t-il pu se former, il y a longtemps que des biologistes faisaient une hypothèse — et des physiciens, ou des chimistes — faisaient une hypothèse très intéressante. Ils employaient eux aussi, bien avant les chaînes de Markov, une comparaison langagière. Ils disaient, si vous vous donnez des [120 :00] lettres au hasard et que vous tirez, vous avez très peu de chances de tomber sur un mot. Mais si vous vous donnez des syllabes, vous avez évidemment beaucoup plus de chances de former un mot. Et cela leur donnait l’idée de : « état intermédiaire ». À savoir, ils disaient : est-ce qu’on ne pourrait pas concevoir l’apparition du vivant sous la forme suivante : à travers un passage par des états intermédiaires à condition que ces états soient stables, et l’on passerait d’état intermédiaire en état intermédiaire par fluctuations, par fluctuations autour de la moyenne de l’état précédent ?
Ça veut dire [121 :00] quoi en cosmobiologie ? Ça veut dire que, par exemple, on part d’un état de corps particulier qu’on appelle les « colloïdes » qui n’ont rien de vivant, les colloïdes, voilà, première état. Dans certaines conditions à déterminer, il y a une fluctuation, ce qu’on appellera une fluctuation par rapport à l’état moyen du colloïde. Et cette fluctuation donne ce qu’on appelle un « coaservat », par exemple, un coaservat de composés, de composés carbonés. Bon, des fluctuations par rapport à un état moyen, on en trouve constamment – remarquez, hein, je le dis tout de suite [122 :00] parce que c’est essentiel — on en trouve constamment. Même dans un gaz, avec répartition brownienne des particules, avec répartition des particules au hasard, vous trouvez constamment des fluctuations. Qu’est-ce qu’on appellera une fluctuation ? Un moment où un nombre insolite de particules se groupe dans une région précise. Mais ces fluctuations sont comme immédiatement neutralisées ou compensées, d’habitude. C’est ça qui empêche l’état intermédiaire de prendre. À peine il est formé qu’il est déjà dissipé.
Donc le problème de la cosmobiologie, ce sera [123 :00] : est-ce qu’on peut imaginer des conditions où une fluctuation se formant par rapport à un état donné, la fluctuation ne serait plus dissipée mais, au contraire, amplifiée de manière à donner un nouvel état ? C’est un problème fondamental. Alors laissons de côté pour le moment le problème des conditions, puisqu’on le retrouvera : à quelles conditions une fluctuation au lieu de s’annuler, s’amplifie ? Mais essayons de mieux comprendre. On part de l’état colloïde, fluctuation par rapport à son état moyen, ça nous donne un nouvel état, coaservat de composés carbonés, un ensemble de composés carbonés. [124 :00] Il y a toutes chances encore une fois pour que la fluctuation s’annule. Supposons qu’elle ne s’annule pas à certaines conditions ; à ce moment-là, bon, d’intermédiaires, il y aura de nouveaux intermédiaires — je saute, c’est pour aller vite — on finira par la possibilité de chaînes d’enzymes, ou de chaînes de protéides, chaînes de protéides qui ne formeront pas encore un organisme vivant.
Mais d’état intermédiaire en état intermédiaire, c’est-à-dire de fluctuation amplifiée en fluctuation amplifiée, est-ce qu’on ne peut pas concevoir d’une autre manière, d’une nouvelle manière le problème de l’origine de la vie ? C’est très [125 :00] différent d’une formation au hasard, vous voyez. C’est quoi ? C’est-ce que je suis en train de vous raconter, c’est une chaîne de Markov mais de l’évolution, c’est une chaîne de Markov. Vous avez exactement une série de tirages dépendants les uns des autres, une série de tirages au hasard, dépendants les uns des autres, où chaque fois la fluctuation marque un hasard qui, s’il n’est pas annulé, s’il a des conditions d’amplification, va aller vers un nouveau tirage qui à nouveau va produire. Vous avez un enchaînement de tirages dépendants les uns des autres, bien que chaque tirage soit aléatoire, vous avez une chaîne de Markov. Ou une espèce de chaîne de Markov. On garde pour le moment… là, il faut, tout ce que je dis ne vaut rien si [126 :00] on oublie le problème : c’est très bien tout ça, mais pour qu’une fluctuation engendre un nouvel état, qui sera dans un rapport semi-aléatoire, vous voyez, il sera exactement dans un rapport aléatoire partiellement dépendant de l’état précédent. Le coaservat de composés carboné, sera par rapport à l’état colloïdal dans un état de, dans un rapport de semi-dépendance, dans un rapport aléatoire semi-dépendant, à condition — encore une fois, je n’ai pas du tout encore posé, répondu à la question — à condition que la fluctuation ne s’annule pas.
Il y a un texte de [Charles] Darwin, une lettre de Darwin, moi que je trouve géniale, où Darwin jeune, non pas si jeune d’ailleurs, il dit : on nous dit tout le temps si la vie avait commencé, pourquoi, pourquoi elle ne pourrait pas [127 :00] recommencer aujourd’hui, hein ? C’était les anti-évolutionnistes du temps de Darwin qui invoquaient déjà cet argument : si ça s’est fait une fois, pourquoi que ça ne se ferait pas une autre fois ? Et Darwin répondait, ben justement, c’est très difficile aujourd’hui qu’une autre forme de vie se forme, disait Darwin. Et pourquoi ? Supposez que dans un petit étang — il est génial, ce texte, génial, parce qu’il nous donne la réponse — supposez que dans un petit étang — à croire que dans une grande mer, ça ne jouerait pas — supposez que dans un petit étang se forment des coaservats, à partir d’états colloïdaux. Eh ben aujourd’hui, ils n’ont aucune chance de faire une fluctuation amplifiante. Pourquoi ? Parce qu’ils seraient mangés tout de suite, [128 :00] ils seraient mangés tout de suite par les vivants déjà là. Ils seraient immédiatement absorbés ; il y aurait toujours un poisson pour les avaler. Alors si la vie, elle n’a pu se former qu’une fois, c’est pour une raison simple : c’est que dès qu’elle s’est produite, elle consomme tout. Alors il n’y a pas beaucoup de risques qu’une autre forme de vie puisse se former avec des chaînes d’un autre type ; les chaînes d’un autre type seraient immédiatement englouties. Mais sa remarque « dans un petit étang », elle doit nous enchanter à nouveau car elle nous met sur la voie de la solution : à quelles conditions une fluctuation peut être amplifiante ? Mais c’est une voie, on ne sait pas encore, on ne sait pas encore.
Voilà la seconde direction de recherche, la cosmobiologie. Or je [129 :00] conclue ce point, encore une fois, parce que je voudrais aller très doucement. Voyez que cet enchaînement d’états stables intermédiaires, où l’on passe de l’un à l’autre par fluctuations amplifiées ou amplifiantes, forment une chaîne de Markov, ou une espèce de chaîne de Markov, c’est-à-dire un enchaînement de phénomènes aléatoires, partiellement dépendants. [Pause]
Troisième ordre de recherche, les recherches faites par [Ilya] Prigogine et Isabelle Stengers. Car c’est justement des recherches portant sur les fluctuations, et posant directement — c’est le chapitre 6 de [130 :00] « La sainte alliance » [au fait, La Nouvelle alliance (Paris : Gallimard, 1979)] — posant directement la question : dans quelles conditions une fluctuation est-elle amplifiante, et dans quelles conditions, au contraire, elle s’annule ou est compensée ? [Pause] Donc, ils devraient nous donner une réponse au problème que nous posions tout à l’heure. [Pause] L’état, l’état primaire où les fluctuations sont immédiatement compensées, c’est encore une fois l’état de la théorie cinétique des gaz. [Pause] Un grand [131 :00] nombre de particules se distribuent dans un milieu fluide, fluide — même à la limite, on dira idéalement fluide — un grand nombre de particules dans un milieu idéalement fluide, qu’est-ce qui se passe pour leur mouvement ? Elles ont, chaque particule a ce qu’on appelle un « libre parcours ». Le libre parcours, c’est quoi ? C’est la distance moyenne parcourue par une particule, avant d’être choquée par une autre particule.
Voyez. J’insiste sur « libre parcours ». Pourquoi il faut le milieu fluide d’un gaz ? Parce que dans un solide, les particules ne sont plus indépendantes. Pourquoi les particules ne sont plus indépendantes dans un solide ou même [132 :00] dans un liquide ? Parce que l’état même du solide ou du liquide les rassemble dans une région [Pause] où elles perdent leur indépendance même ; elles sont prises dans la région du solide de telle manière qu’elles n’ont plus de libre parcours, elles perdent leur libre parcours. [Pause] Dans un gaz, avec mouvement brownien des particules, voyez, le mouvement brownien, c’est donc : une particule fait un parcours jusqu’à ce qu’elle soit, jusqu’à ce qu’elle soit, jusqu’à ce qu’elle soit choquée par une autre particule qui l’envoie dans une autre direction, etc. [133 :00] Et vous avez le type d’une répartition au hasard soumise à des lois de probabilités dans la mesure où vous avez un très grand nombre de particules. Une fluctuation, c’est donc lorsque une grande proportion de ce grand nombre de particules, ou bien une proportion importante, se trouve dans une même région.
Immédiatement, je dis, c’est corrigé, c’est annulé ; pourquoi ? [Pause] À cause, nous disent Prigogine et Stengers, à cause de la diffusion : en effet, en milieu fluide, à cause de la diffusion qui couple toutes les régions du système. [Pause] [134 :00] C’est parce que la diffusion qui couple toutes les régions du système — et notamment qui couple la région fluctuante, c’est-à-dire la région où a apparue la concentration anormale de particules, avec l’environnement [Pause] — c’est parce que la diffusion entre donc la région où a lieu la fluctuation et l’environnement est très, est très élevée que la fluctuation est immédiatement compensée et la répartition probabilitaire des particules rétablie. [Pause] C’est donc la diffusion, bon. [135 :00]
Alors, la réponse à ma question, c’est donc : il faut qu’un seuil soit franchi pour qu’une fluctuation ne soit pas annulée, ne soit pas compensée, mais soit amplifiée. Pour qu’une fluctuation s’amplifie au point d’engendrer un nouvel état, qu’est-ce qu’il faudrait ? [Pause] Il faudrait que la région fluctuante soit, d’une manière ou d’une autre, à l’abri [Pause] [136 :00] de la rapidité de diffusion qui rétablit l’état moyen. [Pause] Prigogine, il va même loin, parce qu’il dit : vous savez, ça vaut pour nos sociétés aussi. Il y a tout le temps, comment expliquer que nos sociétés qui étaient très complexes soient, soient si stables, alors qu’à chaque instant se forment des fluctuations, c’est-à-dire des écarts par rapport à la moyenne ? Eh ben, il dit, c’est qu’il y a un équivalent de la fluidité : c’est la rapidité de l’information. C’est pour ça que la rapidité de l’information, c’est fondamentalement quelque chose de… c’est lié au maintien de l’ordre. [137 :00] C’est la rapidité de l’information qui immédiatement annule les fluctuations, les ramène à, les, les abat.
Et il dit là, dans une phrase intéressante, il dit, ben oui, c’est pour ça que les fluctuations qui risquent de survivre, elles ont très souvent leurs origines dans des groupes marginaux, parce que les groupes marginaux sont protégés contre la rapidité de l’information, de la diffusion. Donc ça se diffusera après, mais justement, [Pause] ce sera une, [Pause] un truc amplifié, là, une — j’ai perdu mon mot, une, ahhh… – une [138 :00] fluctuation amplifiante ou amplifiée. Vous comprenez un petit peu ? Donc voilà la réponse à la question : pour qu’une fluctuation, c’est-à-dire un écart par rapport à un état moyen petit « a » puisse s’amplifier, il faut que, d’une certaine manière, il échappe à la rapidité de la diffusion. À quelles conditions est-ce qu’il peut échapper à la rapidité de la diffusion ? C’est que, si se fait une accumulation — voilà en gros la réponse, oui, je n’en vois même pas d’autre — si se fait une accumulation dans un milieu restreint et que ce milieu restreint soit à l’abri [139 :00] du milieu environnant qui viendrait annuler la fluctuation, il viendrait compenser la fluctuation, c’est-à-dire lui assurer son annulation par rediffusion ; il faut que le milieu restreint soit protégé de la diffusion. [Pause]
Là, en plein, vous avez à nouveau — ça m’étonne qu’il n’y fasse pas référence, je ne crois pas qu’il le fasse même dans le reste — vous avez en plein une chaîne de Markov. Il donne un exemple très curieux, Prigogine ; il donne l’exemple des termitières, qui est typiquement, qui peut être interprété comme une chaîne de Markov. Alors c’est très intéressant parce que la termitière, on peut se dire que la termitière, c’est un tel chef d’œuvre que c’est un système centré, [140 :00] c’est un système arborescent, centré. Non, pas forcément. Il y a une interprétation de la termitière chez…, qui est typiquement chaîne de Markov. À savoir : les termites transportent des petites boules de terre, et puis elles les lâchent en cours de route ; vous me suivez ? Vous êtes un termite, vous transportez des boules de terre, vous les lâchez en cours de route. En les lâchant, les termites les marquent d’une hormone. [Pause] Cette hormone a pour propriété d’attirer d’autres termites qui lâchent alors aussi. Bon, qu’est-ce qu’il faut pour que les piliers, au moins les piliers de la termitière, [141 :00] soient déjà établis ? Il suffit d’un facteur aléatoire, il suffit d’une fluctuation, à savoir que le hasard, l’aléatoire, aléatoirement, un grand nombre de termites aient lâchés leur boule de terre dans une portion restreinte de leur territoire. C’est une fluctuation par rapport à la répartition moyenne des boules de terre. Vous me suivez ? On en aura vu aujourd’hui, les termites, les… hein ?
Ça c’est le facteur aléatoire, qu’un grand nombre, qu’un nombre considérable de termites lâchent leur boule de terre dans une portion restreinte du territoire des termites. Là-dessus, tout s’enchaîne, [Pause] [142 :00] tout s’enchaîne, c’est-à-dire, si ça se fait, ça, vous avez une portion du territoire qui par nature résiste déjà à la diffusion. Pourquoi ça résiste à la diffusion ? À cause de l’hormone, qui attire d’autres termites qui vont poser leur boule de terre aussi. Et vous aurez, à égale distance des piliers qui s’élèveront. Pourquoi égale distance ? La distance sera égale à la portée olfactive de l’hormone. Voilà. Le début, au moins — car il faut, il faut aller très lentement, il faut être prudent — le début de la construction d’une termitière peut être ici assimilé à une chaîne de Markov, [143 :00] c’est-à-dire, une succession de phénomènes aléatoires semi-dépendants. Voyez que le facteur de détermination, c’est l’hormone, le facteur aléatoire, c’était le lâcher au hasard des boules de terre ; la condition pour que la fluctuation s’amplifie, c’était qu’un certain nombre de termites lâchent leur boule de terre dans une périphérie très restreinte, très… [Pause] Ça c’est le troisième.
Quatrième ordre de recherche. [Pause] C’est les recherches d’un cybernéticien [144 :00] aussi, d’un biologiste cybernéticien qui s’appelle Pierre Vendryes, V-E-N-D-R-Y-E-S ; ne le confondez pas avec un autre Vendryes [Joseph], c’était son père et il était linguiste. Et Vendryes, son schéma est très simple, je vous le dis parce qu’il est très beau, en même temps, et puis il va nous faire avancer. Il dit : essayons de définir le vivant par la constitution des milieux intérieurs. L’organisme, c’est une instance qui s’est constituée un milieu intérieur, et plus le milieux intérieur est solide — et on voit ce qu’il veut dire, c’est en plein dans notre problème du coup ; voyez, on retrouve la notion de milieu intérieur maintenant, mais à un autre niveau — [145 :00] à savoir en effet, le milieu intérieur, c’est dans le territoire d’un vivant, ça fait partie du territoire du vivant, et il est d’autant plus consistant qu’il a acquis ses autorégulations, donc il échappe à la fluidité du milieu extérieur, ou il échappe à la diffusion. La constitution d’un milieu intérieur est, à la lettre, une — c’est curieux que j’aie perdu le mot maintenant — une fluctuation amplifiante. [Pause]
Quelle est la conséquence immédiate du milieu intérieur ? La conséquence immédiate du milieu intérieur, c’est que le vivant [146 :00] va entrer ou va pouvoir entrer en relation aléatoire avec le milieu extérieur. Un vivant qui a un milieu intérieur entre par là même en relation aléatoire avec le milieu extérieur et non plus en relation de déterminisme. Pourquoi ? Il faut voir de quel point de vue. Un milieu intérieur implique constitution de réserves ; le vivant a donc des réserves. En tant qu’il a des réserves, il n’a pas besoin d’une vie extérieure. Donc vous voyez, les conditions sont très précises. – [Une étudiante fait une remarque inaudible, puis rit, qui interrompt Deleuze] Merci. — Vous voyez, [147 :00] hein, je ne sais plus ce que vous voyez. [Rires]
Oui, en tant, bien entendu, si le vivant a faim, par exemple, il n’est pas en relation aléatoire avec le milieu extérieur, comprenez. Il a faim, il cherche à manger à l’extérieur. Mais ce n’est pas notre problème ; nous ne considérons pas le vivant en tant qu’il a faim. Nous considérons le vivant en tant qu’il a des réserves, constituées par ses milieux intérieurs. Donc, au moment et dans la mesure et pour autant qu’il a des réserves constituées par son milieu intérieur, il est indépendant du milieu extérieur, tant que durent les réserves ; il en sera à nouveau dépendant, réserves épuisées. J’ai, par exemple, dans mon organisme des sucres, des protides, etc., je peux attendre pour manger ; [148 :00] je suis indépendant de mon milieu extérieur.
Qu’est-ce que je fais alors ? C’est bien connu : je me balade. Je me balade. Qu’est-ce que c’est une balade ? C’est une entrée en relation aléatoire avec le milieu extérieur. [Pause] Si j’ai faim, je ne me balade plus, si mes réserves sont épuisées, c’est fini ça. Enfin je me balade ou je dors. Et si je ne dors pas, ben, je me balade, c’est la flânerie, quoi. Les lions tantôt dorment, tantôt flânent, tantôt se baladent. Qu’est-ce que ça donnera ? En d’autres termes, je suis en relation aléatoire avec le milieu extérieur. Est-ce que c’est… alors qu’est-ce que ça va donner la relation aléatoire du [149 :00] vivant ? L’autonomie du vivant se définit par sa relation aléatoire avec le milieu extérieur. Eh bien, plus Vendryes va très loin, puisqu’il dit, c’est parfait, il dit, il y a une autonomie organique, qu’on vient de définir, mais il y a aussi une autonomie intellectuelle. Elle est vérifiée par l’hypothèse, car l’hypothèse, c’est la relation aléatoire de l’idée avec le monde extérieur. C’est une bonne idée, double autonomie. Ben là, ça concerne le cerveau, le cerveau en tant que milieu intérieur entre dans une relation aléatoire avec le monde extérieur. Cette relation aléatoire est effectuée par ce qu’on appelle les hypothèses. [Pause]
Alors qu’est-ce que ça va être, l’allure du mouvement de la flânerie, de la balade ? [150 :00] Alors, très bizarrement, ça va ressembler à un mouvement brownien, [Pause] mais ça ne va pas être un mouvement brownien. [Pause] Voyez ? Et Vendryes, pour désigner ce type de mouvement … ça ne va pas être un mouvement brownien puisque le mouvement brownien est défini par l’indépendance des particules dans un milieu de diffusion, dans un milieu fluide, idéalement fluide. Là, au contraire, il s’agit de la relation aléatoire d’une particule pourvue d’un milieu intérieur contre-aléatoire. [Pause] [151 :00] Le milieu intérieur est, en effet, contre-aléatoire. [Pause] Vous comprenez un peu ? Je ne sais pas, vous êtes peut-être déjà, peut-être trop fatigués, mais j’en arrive presque au bout.
Vendryes, il était finalement, oui, il dit, regardez, et il prenait comme exemple, il a été l’un des premiers à photographier les mouches ou les moustiques qui volent, le vol des mouches et des moustiques, et c’est évident que c’est un mouvement très proche, qui semble très proche d’un mouvement brownien. Voyez ? Dans des conditions de fluidité idéales, on a l’impression que les mouches, les moustiques se comportent tout à fait en mouvement brownien. Vous avez déjà vu des mouches [152 :00] voler, mais c’est une différence absolue. Il y a une différence absolue encore une fois, puisque, au lieu de particules indépendantes, vous avez un double mouvement, relation aléatoire avec le milieu extérieur en tant qu’elle dépend d’un milieu intérieur contre-aléatoire. Il appellera ça un « mouvement brownoïde ». [Pause] Il ira jusqu’à dire que l’histoire, que l’histoire des hommes est pleine de mouvements brownoïdes.
Et dans un livre un peu bizarre, De la probabilité en histoire [Paris : Albin Michel, 1952], il analysera l’expédition de Bonaparte en Égypte, l’expédition d’Égypte et la fameuse poursuite Napoléon-Nelson en termes brownoïdes, en [153 :00] disant, en effet, que ça a été absolument une chaîne de parcours brownoï… quasi-brownien que Napoléon a fait et que Nelson poursuivant Napoléon a fait, chacun ignorant où l’autre était, et qui parcourent la Méditerranée comme vraiment dans des distributions au hasard et déterminés par les réserves qu’ils ont. Tout à fait, bon, on peut trouver ça en histoire.
Pour aller très, très vite, ça reviendrait à dire, quoi ? Moi ça m’intéresserait beaucoup, ça revient à dire que l’histoire, elle ne cesse pas d’être parcourue par ces mécanismes semi-aléatoires. Ou si vous préférez, [Raymond] Ruyer a une très jolie formule dans son livre La genèse des formes vivantes [Paris : Flammarion, 1958], « l’histoire, ce n’est pas du logos, c’est du jargon ». L’histoire jargonne, et, ouais, c’est du jargon, elle opère : ce n’est pas du hasard, ce n’est pas du déterminisme, [154 :00] ce n’est pas l’idée, c’est-à-dire, ce n’est pas le logos ; c’est des phénomènes aléatoires partiellement dépendants, c’est des chaînes de Markov.
Car ce que, ce que Vendryes appelait, il me semble, d’une manière encore imparfaite, [Pause] « mouvement brownoïde », on a tout intérêt, il me semble, à en faire la théorie systématique sous la forme de chaînes de Markov. Le rapport avec le milieu, le rapport aléatoire avec le milieu sera du type chaîne de Markov, c’est-à-dire phénomène aléatoire partiellement dépendant. Exemple du phénomène aléatoire partiellement dépendant par excellence : la course d’un taxi dans une journée. [155 :00] C’est très proche d’un mouvement brownien, la course d’un taxi dans une journée. Ça a passionné les sociologues ; il y a beaucoup de gens qui se sont occupés de la course d’un taxi dans une journée. Mais vous voyez bien que ce n’est quand même pas exactement brownien, pour une raison très simple : c’est beaucoup plus un enchaînement de phénomènes aléatoires partiellement dépendants puisque chaque course dépend partiellement de la course précédente, ne serait-ce que du point de vue de la prise en charge. Si bien que la course d’un taxi dans une journée est beaucoup plus proche d’une chaîne de Markov que d’un mouvement brownien. C’est ce que Vendryes essayait de dire en parlant de mouvements brownoïdes et non pas browniens.
Bien alors, ce qui revient à dire quoi, tout ça ? Ben, [156 :00] que d’une certaine manière, voilà, on tient peut-être notre solution. Je veux dire, tout ça, qu’est-ce que ça nous amène ? Les chaînes de Markov, s’il fallait essayer d’en donner une formule alors pas scientifique, une formule philosophique, je dirais : c’est un ensemble de ré-enchaînements sans enchaînement préalable. Et ça alors du coup, j’y tiens ; tout le reste, je m’en fous, c’est pour arriver à ça. C’est pour arriver à cette chose qui pour moi fait mystère : comment est-il possible qu’il y ait des ré-enchaînements s’il n’y a pas d’enchaînement ? Supposez que des enchaînements se fassent au hasard, mais les ré-enchaînements ne sont pas au hasard. Voilà, je dirais une chaîne de Markov, c’est ça. [157 :00] Il n’y a plus d’enchaînement, il n’y a plus que des ré-enchaînements, c’est-à-dire des thèmes indépendants l’un de l’autre se ré-enchaînent, au lieu qu’il y ait enchaînement de thèmes dépendants. Au lieu qu’il y ait enchaînement de thèmes dépendants, il y a ré-enchaînement de thèmes indépendants, il y a un primat du ré-enchaînement sur l’enchaînement – ce n’est pas la première fois [Pause] — et les chaînes de Markov nous donnent la structure mathématique d’un tel événement.
Alors ça m’apporte beaucoup parce que c’est ma réponse à : comment se ferait la transmission dans le cerveau ? La transmission dans le cerveau se fait par ré-enchaînement [158 :00] en séries indépendantes. Ce serait beaucoup plus proche des chaînes de Markov — d’où l’idée d’un cerveau probabilitaire — beaucoup plus proche d’une chaîne de Markov que d’une transmission électrique. Mais alors, on tient tout. Parce que je passe au dernier aspect que j’annonce juste, qu’est-ce que ça veut dire tout ça pour le cinéma ? Est-ce qu’on ne pourra pas dire aussi, il y a deux régimes de l’image très différents de ce point de vue-là ? Il y a eu un régime de l’enchaînement d’images qui a inspiré la conception classique du montage, et c’était un enchaînement sensorimoteur des images. [Pause] [159 :00] Mais est-ce que le cinéma dit moderne ne nous proposerait pas des structures très différentes qui serait des ré-enchaînements d’images indépendantes ? Il n’y aurait plus d’enchaînement, il n’y aurait que des ré-enchaînements. [Fin de la séance] [2 :39 :18]
For archival purposes, the augmented and new time stamped version of the complete transcription was completed in August 2021. Additional revisions were added in February 2025 and June 2025. The translation was completed in January 2025, published in August 2025.